Mostrando artículos por etiqueta: bachillerato

Título: Los números complejos (ed. 2023)
Sección: iCartesiLibri
Bloque: Álgebra
Unidad: Números y operaciones
Nivel/Edad: Bachillerato y Universidad (16 años o más)
Idioma: Castellano
Autora: María José García Cebrian
ISBN: 978-958-56476-0-2
![]() Haz clic aquí para ver una versión en pdf
Haz clic aquí para ver una versión en pdf
![]() Haz clic en la imagen para abrir el recurso
Haz clic en la imagen para abrir el recurso
Puedes encontrar todos los libros interactivos de iCartesiLibri en
https://proyectodescartes.org/iCartesiLibri/index.htm - Ver Créditos
Este material está publicado bajo una licencia:

Creative Commons Reconocimiento-NoComercial-CompartirIgual 4.0 Internacional

Título: Funciones
Sección: iCartesiLibri
Bloque: Análisis matemático
Unidad: Funciones
Nivel/Edad: Bachillerato y Universidad (16 años o más)
Idioma: Castellano
Autores: Norma Patricia Apodaca Alvarez
ISBN: 978-84-18834-56-1
 versión en pdf con enlace a los interactivos
versión en pdf con enlace a los interactivos
![]() Haz clic en la imagen para abrir el recurso
Haz clic en la imagen para abrir el recurso
Puedes encontrar todos los libros interactivos de iCartesiLibri en
https://proyectodescartes.org/iCartesiLibri/index.htm - Ver Créditos
Este material está publicado bajo una licencia:
Creative Commons Reconocimiento-NoComercial-CompartirIgual 4.0 Internacional

Título: Cuarta revolución industrial. Fundamentos.
Sección: iCartesiLibri
Bloque: Ingeniería
Unidad: Fundamentos de tecnología
Nivel/Edad: Bachillerato y universidad (16 años o más)
Idioma: Castellano
Autores: Juan Guillermo Rivera Berrío y Ramiro Antonio Lopera Sánchez
ISBN: 978-84-18834-53-0
![]() Haz clic aquí para ver una versión en pdf
Haz clic aquí para ver una versión en pdf
![]() Haz clic en la imagen para abrir el recurso
Haz clic en la imagen para abrir el recurso
Puedes encontrar todos los libros interactivos de iCartesiLibri en
https://proyectodescartes.org/iCartesiLibri/index.htm - Ver Créditos
Este material está publicado bajo una licencia:
Creative Commons Reconocimiento-NoComercial-CompartirIgual 4.0 Internacional
Estadística, probabilidad e inferencia es un libro interactivo para bachillerato que pertenece al Proyecto iCartesiLibri de la RED Descartes.
Este libro consta de una serie de capítulos que tratan temas de estadística, combinatoria, probabilidad e inferencia. En cada capítulo además de las definiciones, conceptos y fórmulas introducidos, se presentan vídeos, actividades interactivas de introducción y para practicar, autoevaluaciones y ejercicios resueltos.
Capítulos del libro:
Introducción a la estadística, definiciones, tablas de frecuencia y diagramas estadísticos y medidas de centralización y dispersión.
Variable estadística bidimensional, elaboración de tablas y gráficos, correlación y regresión. Ejercicios resueltos.
Recuento de datos, variaciones, permutaciones y combinaciones sin y con repetición. Criterios para aplicar las técnicas de recuento.
Introducción, espacio muestral, tipos de sucesos, probabilidad, probabilidad condicionada y teorema de Bayes.
5. Variable estadística discreta
Función de probabilidad, distribución binomial, otras distribuciones: hipergeométrica, de Poisson, geométrica, binomial negativa y uniforme.
La distribución Normal, Manejo de la tabla N(0,1), Manejo inverso de la tabla N(0,1). Aproximación de una binomial por una normal.
7. Inferencia estadística. Muestreo
Estudio por muestreo, distribución en el muestreo de la proporción y distribución en el muestreo de las medias muestrales. Teorema central del límite.
8. Inferencia estadística. Intervalos de confianza
Intervalo de confianza, estimación puntual y estimación por intervalos. Error máximo admisible y tamaños muestrales.
Hipótesis nula y alternativa, planteamiento general de un problema de contraste y tipos de errores.

Título: Matemáticas para las Ciencias 3
Sección: iCartesiLibri
Bloque: Análisis matemático
Unidad: Integración y Cálculo vectorial
Nivel/Edad: Universidad (18 años o más)
Idioma: Castellano
Autor: Marco Iván Olea Olvera
ISBN: 978-84-18834-51-6
![]() Haz clic aquí para ver una versión en pdf
Haz clic aquí para ver una versión en pdf
![]() Haz clic en la imagen para abrir el recurso
Haz clic en la imagen para abrir el recurso
Puedes encontrar todos los libros interactivos de iCartesiLibri en
https://proyectodescartes.org/iCartesiLibri/index.htm - Ver Créditos
Este material está publicado bajo una licencia:
Creative Commons Reconocimiento-NoComercial-CompartirIgual 4.0 Internacional

Prometeo es un subproyecto de la Red Educativa Digital Descartes que contiene unidades didácticas interactivas, principalmente de matemáticas y física, creadas con Descartes. Los materiales cubren diferentes niveles educativos y forman parte del proyecto Prometeo del Instituto de Matemáticas de la UNAM (Universidad Nacional Autónoma de México).
Los recursos están agrupados a su vez en diferentes proyectos, a los cuales se accede desde la página web del subproyecto. En cada una de las unidades se puede consultar la información sobre su contenido, visualizar el recurso o bien descargarlo para su uso en local.
Unidades didácticas interactivas para su funcionamiento en celulares
Adaptación de unidades interactivas para su funcionamiento en dispositivos móviles. Consiste en 21 unidades variadas de matemáticas para licenciatura. Cada escena incluye una versión para computadora y otra específica para móviles. Incluye actividades de:
-
- Álgebra: Números y operaciones, Ecuaciones algebraicas, Relaciones Algebraicas, Álgebra Lineal y Lógica.
- Análisis: Límite y continuidad de funciones, Derivación de funciones y Representación gráfica de funciones.
- Geometría: Geometría Analítica Tridimensional.
Campos Vectoriales y Ecuaciones de Maxwell
Comprende cuatro unidades que abordan los campos vectoriales, el cálculo vectorial y su aplicación en las ecuaciones del electromagnetismo de Maxwell. El nivel al que están orientadas es introductorio para licenciatura, pero pueden ser usadas también a nivel bachillerato.
Recursos educativos interactivos de matemáticas para el bachillerato
Abarca los diversos contenidos del área de matemáticas que se contemplan en el Bachillerato de México, o preparatoria para el acceso a la universidad. Comprende tres cursos, de primer a tercer grado. Estos cursos se corresponden en el sistema educativo español con 4º de educación secundaria obligatoria (ESO) y 1º y 2º de Bachillerato. Los materiales se clasifican atendiendo a las siguientes disciplinas:
-
- Aritmética.
- Álgebra.
- Geometría: Geometría Plana, Trigonometría y Geometría Analítica Plana.
- Cálculo: Funciones y sus gráficas y Cálculo diferencial e integral.
Unidades Didácticas Interactivas de Introducción al Pensamiento Matemático
En este proyecto se encuentran recursos digitales interactivos de geometría, álgebra y análisis.
-
- Geometría: Geometría métrica plana y Geometría analítica plana.
- Álgebra: Combinatoria, Grafos, Redes y Números y operaciones.
- Análisis: Sucesiones y progresiones.
Unidades Didácticas Interactivas para la asignatura Taller de Matemáticas
Cada unidad abarca un tema distinto de aritmética, álgebra, trigonometría, geometría y geometría analítica.
-
- Álgebra: Números y operaciones, Expresiones algebraicas, Ecuaciones y sistemas e Inecuaciones y sistemas.
- Análisis: Funciones elementales.
- Geometría: Geometría analítica plana, Geometría plana y Trigonomería.
En el siguiente vídeo se presentan los diferentes proyectos y se muestran, a modo de ejemplo, algunas de las unidades:

Título: Electrónica Digital. Conceptos y Métodos Básicos.
Sección: iCartesiLibri
Bloque: Ingeniería
Unidad: Electrónica
Nivel/Edad: Bachillerato y universidad (16 años o más)
Idioma: Castellano
Autores: Oscar Ignacio Botero Henao
ISBN: 978-84-18834-24-0
![]() Haz clic aquí para ver una versión en pdf
Haz clic aquí para ver una versión en pdf
![]() Haz clic en la imagen para abrir el recurso
Haz clic en la imagen para abrir el recurso
Puedes encontrar todos los libros interactivos de iCartesiLibri en
https://proyectodescartes.org/iCartesiLibri/index.htm - Ver Créditos
Este material está publicado bajo una licencia:
Creative Commons Reconocimiento-NoComercial-CompartirIgual 4.0 Internacional

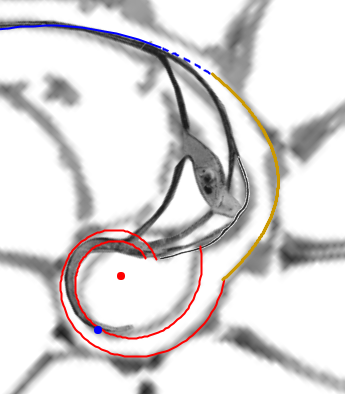
En el primer verticilo del Nautilus encontramos algunos sitios que podemos catalogar como singulares, pues se muestran como específicos respecto a su entorno o que aparentemente se salen de la uniformidad cordobesa que hemos ido detectando y desgranando en los artículos publicados anteriormente sobre la "Ontogenia matemática del Nautilus". Pero, hasta en estas situaciones, la proporción cordobesa y la espiral cordobesa aparece sin más que rascar matemáticamente en la concha del Nautilus, por supuesto con gran mimo.
Esos sitios singulares son:
- La concha embrionaria.
- El sifúnculo en la segunda cámara septal y su relación con la ampliación del fragmacono en el primer septo.
- La confluencia de la pared dorsal con la concha embrionaria en la fase de transición entre el primer y segundo verticilo (cámaras octava, novena y décima).
 |
 |
 |
| La concha embrionaria | El sifúnculo en la segunda cámara septal y la ampliación del fragmacono |
La pared dorsal en la transición entre el primer y segundo verticilo |
En este artículo analizamos estas singularidades y encuadramos su modelado en el contexto cordobés del Nautilus. Para ello, acudimos y aplicamos la invariante enunciada desde el inicio y que reiteradamente hemos ido aplicando en toda la modelación: "Todo punto interior a la concha o sobre ella se obtiene como la intersección de dos espirales cordobesas, una longitudinal similar a la ventral y otra transversal similar a la septal". La siguiente escena interactiva ha servido de base para el análisis de estos sitios singulares.
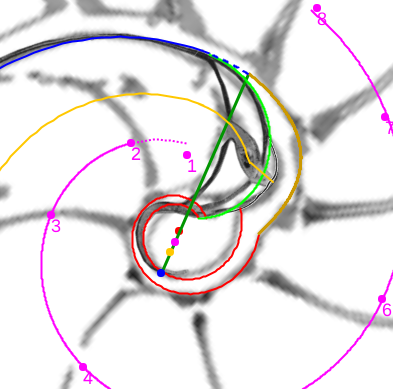
Modelando los sitios singulares del Nautilus en el primer verticilo
Pulsad sobre la imagen para acceder a la escena
En el siguiente pdf (o desde este enlace) tienen desarrollados los contenidos de este artículo
Ontogenia matemática del Nautilus VII
Aquí, hemos dado continuidad a los artículos anteriores (I, II, III, IV, V y VI) y por fin, creo, he cumplido mis deberes, si bien no seré yo quien excluya la posibilidad de que puedan surgir nuevas cuestiones, pues eso es lo que ha ido aconteciendo a lo largo de estos meses en los que les he ido relatando mi investigación a través de este blog de RED Descartes. Pero, independientemente de que afloren nuevas cuestiones o no, sí les indico que éste no será este el último artículo de esta serie, tengo que tratar de completarla adecuadamente y, por tanto, estimo necesario recopilar todo lo expuesto en una escena interactiva de Descartes o en una animación o en un gif animado o en un recurso similar o... y así mostrar la ontogenia matemática del Nautilus en un hilo temporal que recoja su crecimiento, si bien real y paradójicamente éste ha de ser atemporal porque en la literatura existente los datos relativos a los tiempos en el crecimiento de la concha son muy genéricos o yo no he logrado localizarlos. En la siguiente animación reflejamos el modelo uniforme del Nautilus y próximamente podremos aportar el modelo ontogénico (de nuevo les pido un poquito más de paciencia, no nos demos un atracón que pueda conducir a una indigestión).

Modelo uniforme del Nautilus
Y también tendremos que desarrollar alguna cosita en 3D ¿no les parece?... Hasta pronto...

Este obra está bajo una licencia de Creative Commons Reconocimiento-NoComercial-SinObraDerivada 4.0 Internacional
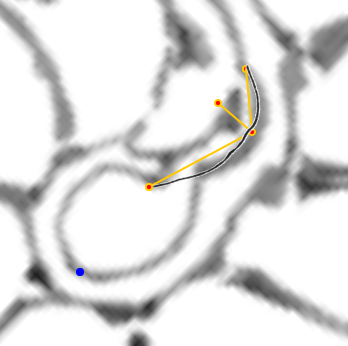
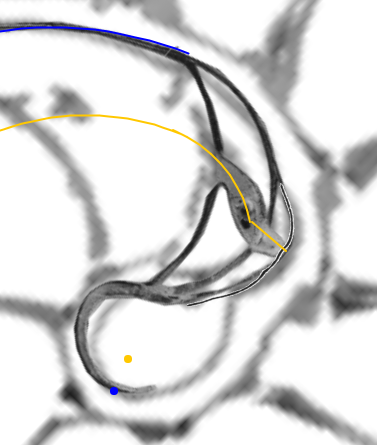
En este nuevo artículo sobre la ontogenia matemática del Nautilus, después de haber modelizado los septos en el segundo y tercer verticilio bajo el invariante de tangencialidad, nos adentramos en la modelización de los septos en el primer verticilo. Esta primera fase de crecimiento vimos que se muestra diversa y con apariencia poco regular, cambiante (menor número de cámaras septales, ocho frente a las dieciséis de la etapa juvenil y adulta, con secciones y amplitudes que cambian como necesidad biológica para alcanzar la flotabilidad) y, ahora, ha llegado el momento de mostrar el modelo matemático que da explicación a esta etapa e introduce la regularidad esperada que parecía no acaecer, pero que queda al descubierto bajo la perspectiva matemática. De nuevo, el hecho de que intervengan dos espirales con diferente polo, en este caso la espiral de la pared ventral y la espiral de los polos de los septos, conduce a proporciones variables entre los radios vectores y consecuentemente a que se formen septos con factores de escala variables. Ello nos conduce y permite determinar las ecuaciones de los septos, los puntos de tangencia con la pared ventral y los de intersección con la pared dorsal (para ello necesitaremos introducir un grosor en el modelo matemático de esa pared, que es lo que físicamente acontece).
Completaremos el contenido de este documento agrupando y relacionando entre sí diferentes puntos que se han ido detectando en este análisis. Unos que denominamos notables, porque matemáticamente son los que establecen el modelo matemático y dan explicación causal al mismo, y que son polos de diferentes espirales. Y otros que catalogaremos como destacables, posible fuente de inspiración matemática futura, y que son centros desde los que algunos objetos se observan con perspectiva angular constante.
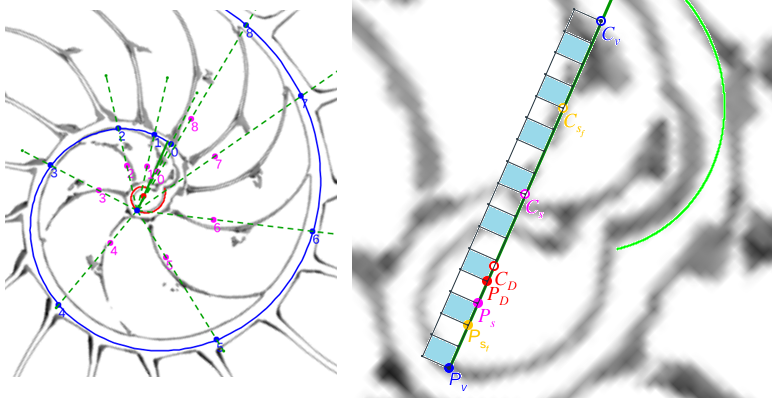 |
|
| Propociones entre los radios vectores de la espiral ventral y los de la espiral de los polos de los septos | Puntos notables y destacables |
Así pues, doy continuidad a los artículos anteriores (I, II, III, IV y V), con un contenido adicional que espero sea de su interés —¡para mí es siempre una satisfacción! ir pudiendo relatarles progresivamente lo que, poco a poco, me cuenta la concha de este animalito—, y he de adelantarles que serán necesarios algunos artículos adicionales porque aún nos quedan secretos que dilucidar en esta ontogenia, en particular lo que acontece en la transición de la fase embrionaria (primera y segunda cámara septal) donde el sifúnculo cambia abruptamente de posición, y en la fase de transición entre el primer y segundo verticilo (cámaras octava, novena y décima) donde al finalizar la primera vuelta se produce el encuentro del fragmacono con la concha embrionaria. Y también habrá que abordar la síntesis o resumen final, es decir, plasmar y reproducir ese modelo ontogénico de la concha del Nautilus.
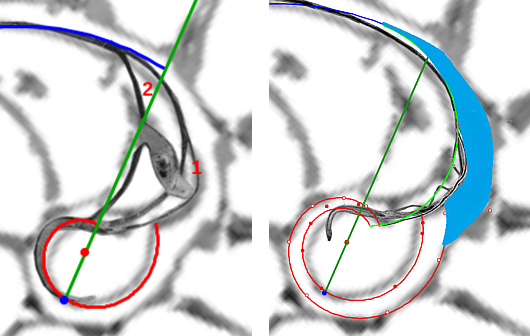 |
|
| El sifúnculo en la segunda cámara septal | Transición entre el pimer y segundo verticilo |
Como observamos, una mirada atenta y un continuo deseo de comprensión nos hace ir visualizando cada vez más detalles que inicialmente pueden parecer nimios, pero que finalmente se han ido mostrando como retos cuya resolución es de interés. Todo ello, a costa de que a ustedes a lo mejor les ocurra como a mi sobrina nieta (Aurora, cerca de los cuatro años) que ayer, al verme una vez más delante de la pantalla de mi ordenador, indagando la imagen de la sección del Nautilus con diversos objetos matemáticos superpuestos, la cual ya ha observado en multitud de ocasiones y quizás hayan sido demasiadas para ella, dijera: "¡Tita!, ¡el tito todavía no ha hecho sus deberes!". Por tanto, espero poder ir finalizando mis deberes, que realmente no son más que satisfacciones aunque requieran esfuerzo y dedicación, y que en el trancurso hacia su final les pueda tener como lectores y juntos podamos desarrollar nuestra vocación como μαθηματικός (mathēmatikós) o amantes del conocimiento.
En el siguiente pdf (o desde este enlace) tienen desarrollados los contenidos de este artículo
Ontogenia matemática del Nautilus VI
Este obra está bajo una licencia de Creative Commons Reconocimiento-NoComercial-SinObraDerivada 4.0 Internacional
.png "Haz click para abrir el recurso")
Título: Curvas y superficies paramétricas (segunda edición)
Sección: iCartesiLibri
Bloque: Geometría
Unidad: Geometría plana y tridimensional
Nivel/Edad: Bachillerato y Universidad (16 años o más)
Idioma: Castellano
Autores: Juan Guillermo Rivera Berrío y Josep Maria Navarro Canut
ISBN: 978-84-18834-35-6
versión en pdf con enlace a los interactivos
![]() Haz clic en la imagen para abrir el recurso
Haz clic en la imagen para abrir el recurso
Puedes encontrar todos los libros interactivos de iCartesiLibri en
https://proyectodescartes.org/iCartesiLibri/index.htm - Ver Créditos
Este material está publicado bajo una licencia:
Creative Commons Reconocimiento-NoComercial-CompartirIgual 4.0 Internacional
Módulo de Búsqueda

Acceso
Lo último
- Séptimo número de la "Revista Digital RED Descartes"
- Publicado el libro interactivo "Gamificando con juegos AJDA"
- Introducción a conceptos estadísticos con DescartesJS: análisis de las Unidades de Media y Mediana del Proyecto @prende
- Anuncio de próxima publicación del libro "Gamificando con juegos AJDA"
- Física y Química. Proyecto ed@d
Lo más leído de lo publicado hace un mes
 CONTACTO
CONTACTO
Canal Youtube

Calculadora Descartes

Versión 3.1 con estadística bidimensional
![]() Código para embeber
Código para embeber
Últimos Comentarios
-
 Excelente contribución a la educación global. Felicitaciones a los organizadores…
Escrito por Ageleo Justiniano Tucto
en %PM, %20 %503 %2023 %13:%Oct
Participantes de tres continentes en el curso para el diseño de libros interactivos
(Difusión)
Excelente contribución a la educación global. Felicitaciones a los organizadores…
Escrito por Ageleo Justiniano Tucto
en %PM, %20 %503 %2023 %13:%Oct
Participantes de tres continentes en el curso para el diseño de libros interactivos
(Difusión)
-
 Estimado Javier Arturo: Agradecemos su reconocimiento al programa de Educación…
Escrito por José Antonio Salgueiro González
en %PM, %22 %458 %2023 %12:%Sep
Abierto el plazo de inscripción en la V Edición del Curso para el Diseño de Libros Interactivos
(Difusión)
Estimado Javier Arturo: Agradecemos su reconocimiento al programa de Educación…
Escrito por José Antonio Salgueiro González
en %PM, %22 %458 %2023 %12:%Sep
Abierto el plazo de inscripción en la V Edición del Curso para el Diseño de Libros Interactivos
(Difusión)
-
Me parece una gran iniciativa en favor de la educación,…
Escrito por JAVIER ARTURO MARTINEZ FARFAN
en %AM, %22 %189 %2023 %05:%Sep
Abierto el plazo de inscripción en la V Edición del Curso para el Diseño de Libros Interactivos
(Difusión)
-
Ildefonso era un hombre de edad y motivaciones educativas similares…
Escrito por José Luis San Emeterio
en %PM, %05 %805 %2023 %20:%Ago
Ildefonso Fernández Trujillo, in memoriam
(Difusión)
-
Yo conocí la fórmula más bella de las matematicas como…
Escrito por Pepin
en %PM, %17 %576 %2023 %14:%Jul
Cálculo diferencial e integral, módulo I
(iCartesiLibri)