Mostrando artículos por etiqueta: bachillerato

En el artículo anterior (Ontogenia III) mostramos la modelización del labio dorsal y del sifúnculo en el primer verticilo del Nautilus y observamos las diferencias que acontecen con respecto a etapas vitales posteriores. En éste, procederemos a realizar un análisis de los tabiques y de las cámaras septales en la primera etapa de la ontogenia del Nautilus y buscaremos caracterizar las particularidades que, de manera evidente, se muestran en ese primer verticilo. Con una simple, pero atenta mirada, se observa que en esa primera vuelta hay un número menor de cámaras que las que se contabilizan en la segunda y tercera, se visualiza una amplitud angular de las mismas que aparece como no uniforme, se intuye un tamaño o capacidad volumétrica diversa con unas cámaras pequeñas y otras grandes con un crecimiento quizás no gnomónico e incluso en algunos casos decreciendo. Muchos detalles en los que aquí profundizamos y sobre los que desvelamos y aportamos explicaciones y posibles causas que lo ocasionan, siempre desde una perspectiva y visión matemática, pero con soporte y referencias biológicas. Le animamos a profundizar en este estudio y a continuar avanzando en la compresión de la esencia nautílica y en el descubrimiento de su belleza oculta.
Los septos en el primer verticilo
Para el análisis de los septos en el primer verticilo nuestra referencia inicial ha de situarse en el modelo uniforme (Galo et al., 2016) donde las cámaras septales se distribuyen siguiendo una amplitud angular de 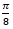 , lo que conduce en cada vuelta a un número de dieciséis tabiques y estos son arcos de espirales cordobesas de ecuación:
, lo que conduce en cada vuelta a un número de dieciséis tabiques y estos son arcos de espirales cordobesas de ecuación:
 (11)
(11)
cuyos polos ![]() están ubicados, a su vez, en otra espiral cordobesa. Ésta última, en ese modelo, se mostraba como la espiral intermedia entre el sifúnculo y la pared dorsal:
están ubicados, a su vez, en otra espiral cordobesa. Ésta última, en ese modelo, se mostraba como la espiral intermedia entre el sifúnculo y la pared dorsal:
![]() (12)
(12)
donde
![]() (13)
(13)
o bien, aproximadamente es la espiral formada por los puntos medios de los radios vectores de la espiral correspondiente a la pared ventral de la concha que, obviamente, se corresponde con el factor 0,5.
Lo antes descrito ha de ser la base primordial sobre la que tenemos que apoyarnos para tratar de canalizar ese análisis y para poder focalizar adecuadamente la búsqueda y para lograr recopilar cuáles son los datos más certeros que nos conduzcan a la compresión del comportamiento y también a su modelación. O al menos a esta última, pues aun teniendo el modelo no siempre es factible saber la causa o posicionarse tajantemente en ella.
Sobre el menor número de cámaras septales en el primer verticilo
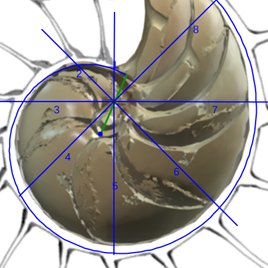
Si observamos las cámaras septales del primer verticilo es evidente, basta hacer un recuento, que el número de éstas son ocho y ello se correspondería con una amplitud angular media de ![]() . No obstante, a primera vista o aparentemente, parece más que esa amplitud se manifiesta como variable y que no es obvio fijarla a priori. Esa cantidad de cámaras es inferior a las existentes en el segundo verticilo en el que son dieciséis con una amplitud constante de
. No obstante, a primera vista o aparentemente, parece más que esa amplitud se manifiesta como variable y que no es obvio fijarla a priori. Esa cantidad de cámaras es inferior a las existentes en el segundo verticilo en el que son dieciséis con una amplitud constante de ![]() y también a las del tercero donde hay un número variable, según el ejemplar y nivel de madurez, pero manteniendo también esa amplitud de .
y también a las del tercero donde hay un número variable, según el ejemplar y nivel de madurez, pero manteniendo también esa amplitud de .
¿Por qué el número de cámaras es menor en el primer verticilo? Ilustrémonos en el devenir de esta etapa inicial del Nautilus acudiendo a algunas referencias descriptivas biológicas y acompañémoslas de una primeras reflexiones matemáticas personales:
- Landman et al. (1989, pp. 12-13) indican que, en la parte correspondiente al labio dorsal, el tercer septo se apoya parcialmente en el segundo e igualmente el segundo en el primero. Por tanto, se induce que el labio dorsal no tiene la suficiente longitud para poder aportar un espacio disjunto para cada septo. Y si eso acontece con la amplitud media interseptal de
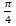 , antes indicada, podemos imaginar el colapso o imposibilidad constructiva a la que se vería abocado el Nautilus si la amplitud fuera aún menor, en concreto la mitad: . Esto lo podemos evidenciar sin más que calcular la longitud del labio dorsal aplicando que la longitud de un arco de la espiral
, antes indicada, podemos imaginar el colapso o imposibilidad constructiva a la que se vería abocado el Nautilus si la amplitud fuera aún menor, en concreto la mitad: . Esto lo podemos evidenciar sin más que calcular la longitud del labio dorsal aplicando que la longitud de un arco de la espiral 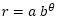 para
para 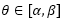 viene dada por:
viene dada por:
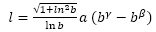 (14) (Galo et al, 2016)
(14) (Galo et al, 2016)y, por tanto, aplicándolo a la ecuación del labio dorsal (3)-(4) y considerando la escala real de las imágenes de la concha del Nautilus que estamos usando en las escenas interactivas, se obtiene que
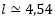 mm. Esto conduce a que en el labio dorsal, para las ocho cámaras que se visualizan en el primer verticilo, hay un espacio medio[1] para cada cámara de unos 0,57 mm lo cual de por sí ya es bastante ajustado, pues en las figuras 13 y 14 procedentes de Ladman et al. y usando la regla de la escena interactiva 2 puede observarse que el apoyo dorsal de cada uno de los tres primeros septos ocupa ese espacio e incluso algo más y de ahí que tengan que superponerse, y que en el resto de las cámaras septales de ese verticilo la distancia entre septos es sólo algo superior. En el supuesto de considerar dieciséis cámaras estaríamos hablando de un espacio medio de 0,285 mm… al Nautilus no le salen las cuentas, ni le trae cuenta[2] elaborar tanto septo, más si consideramos adicionalmente lo que detallamos a continuación.
mm. Esto conduce a que en el labio dorsal, para las ocho cámaras que se visualizan en el primer verticilo, hay un espacio medio[1] para cada cámara de unos 0,57 mm lo cual de por sí ya es bastante ajustado, pues en las figuras 13 y 14 procedentes de Ladman et al. y usando la regla de la escena interactiva 2 puede observarse que el apoyo dorsal de cada uno de los tres primeros septos ocupa ese espacio e incluso algo más y de ahí que tengan que superponerse, y que en el resto de las cámaras septales de ese verticilo la distancia entre septos es sólo algo superior. En el supuesto de considerar dieciséis cámaras estaríamos hablando de un espacio medio de 0,285 mm… al Nautilus no le salen las cuentas, ni le trae cuenta[2] elaborar tanto septo, más si consideramos adicionalmente lo que detallamos a continuación. - Greenwald y Ward (2010) indican que un Nautilus, que pesa en el aire un kg, en el interior del agua en la que habita pesa solo unos pocos gramos, es decir, ponen de manifiesto una de sus características vitales esenciales: su flotabilidad casi neutra. Adicionalmente, detallan como esta flotabilidad se alcanza mediante el vaciado del líquido de las cámaras y cómo éste ha de ir compensándose adecuadamente con el aumento de peso que aporta cada nueva pared septal y también con el correspondiente a la ampliación de la concha ventral que da forma a la cámara habitacional y que es la que pasa a ocupar el animal. El propio animal suma su incremento de peso al crecer (ver figura 26). Y, de nuevo, Landman et al. (1989, p. 15) indican que en la fase de construcción de los primeros septos la proporción entre el volumen de las cámaras septales y el del fragmacono es pequeña y por tanto en esa etapa inicial el animal no alcanza la flotabilidad.
Consecuentemente, en una somera visión, parece fácil concluir que principalmente en la etapa inicial y también en general, ha de ser un empeño básico el lograr economizar en la cantidad de material calcáreo a utilizar ―dato que de nuevo incide y aporta cierto porqué relativo al menor número de septos― y que lo que se use ha de ser compensado por el empuje ocasionado por el volumen de líquido septal que se desaloje, es decir, ha de estar equilibrado con el volumen que alcancen las cavidades septales. Así pues, dado que el Nautilus inicialmente no flota y después sí, se deduce que debe haber una variabilidad volumétrica en las primeras cámaras y que este posible hecho puede ir también relacionado con la amplitud angular interseptos que parece ser variable, según indicamos con antelación. Es necesario y procede analizar esta capacidad de las cámaras, pero en este estudio bidimensional acudiremos sólo al cálculo de la superficie ocupada por las secciones de las cámaras y analizaremos su variación[3]. No obstante, avancemos sin precipitación, que progresivamente van apareciendo diversas facetas y es necesario asentar estas ideas con un material de apoyo adecuado y procede darle el necesario sustento matemático.

Fig. 26. Formación de una cámara y vaciado del líquido de las cámaras que acontece en la cámara anterior. Fuente: Buoyancy in Nautilus, Greenwald L., Ward P.D. (2010).
Sobre la amplitud angular de las cámaras septales en el primer verticilo y en los dos restantes
Ya hemos indicado, e incluso reiterado, que las dieciséis cámaras del segundo verticilo conducen a una amplitud angular entre cámaras de , la cual se mantiene para los septos de tercer verticilo, y que las ocho del primero llevan a una amplitud de . Si consideramos el retardo de la espiral dorsal respecto a la ventral en el segundo y tercer verticilo que es de 2π, al efectuar el cociente 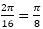 obtenemos el valor numérico antes indicado, pero si aplicamos igual proporción en el primer verticilo ¿qué ocurre? El retardo entre el labio dorsal y la pared ventral en este caso es de 4π y resulta que
obtenemos el valor numérico antes indicado, pero si aplicamos igual proporción en el primer verticilo ¿qué ocurre? El retardo entre el labio dorsal y la pared ventral en este caso es de 4π y resulta que 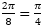 , es decir, un paso entre septos de , paso que casualmente conduciría a los ocho septos observados en ese verticilo[4].
, es decir, un paso entre septos de , paso que casualmente conduciría a los ocho septos observados en ese verticilo[4].
Sí, parece como si estuviéramos enfrascados en el reiterado y recurrente dilema de ¿qué fue antes: el huevo o la gallina?, pero más bien sería una estrategia de observación múltiple, desde puntos de vista opuestos o diferentes, buscando la generación de algún destello que, al menos, difumine un poquito las sombras platónicas en las que toda investigación siempre anda envuelta. Para tratar de provocar la necesaria chispa detonadora vamos a usar la escena interactiva 4.
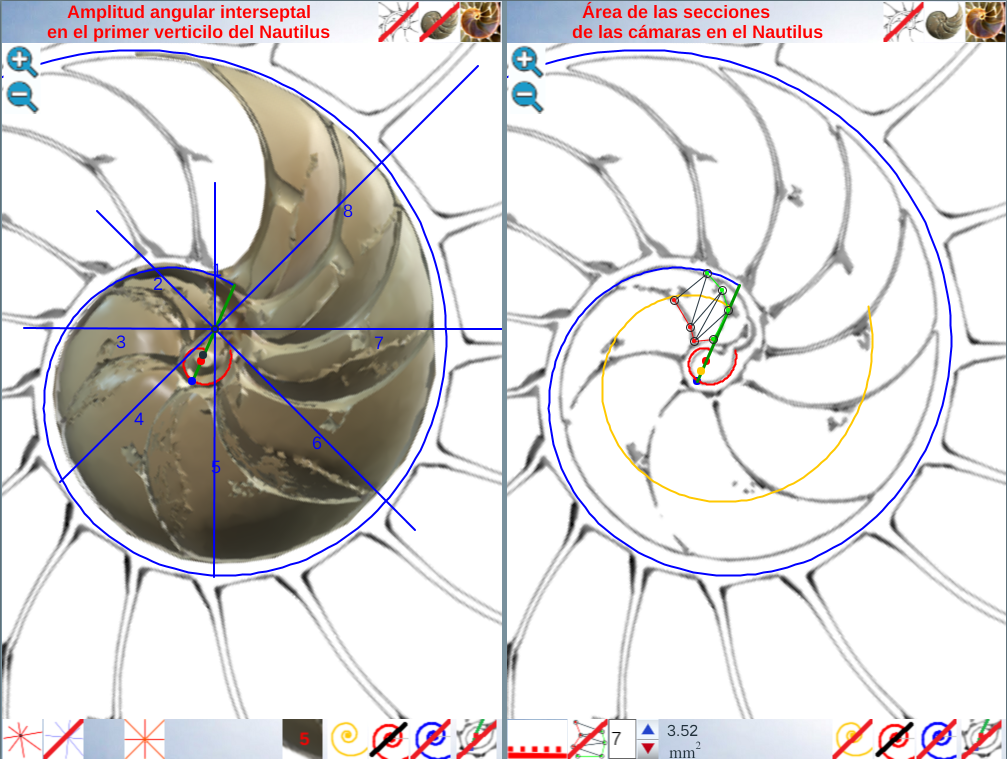
Escena interactiva 4. Amplitud angular interseptal en el primer verticilo y área de las secciones de las cámaras del Nautilus.
Pulse sobre la imagen para interactuar libremente con ella. Si posiciona el ratón sobre los botones tendrá una breve información o consulte las instrucciones.
En el espacio izquierdo de la escena interactiva 4 contamos con herramientas auxiliares que nos permiten analizar la distribución que pueda acaecer en la amplitud septal. Con ellas hemos obtenido las imágenes mostradas en las figuras 27 a 32, que sintetizan el análisis que detallamos a continuación:
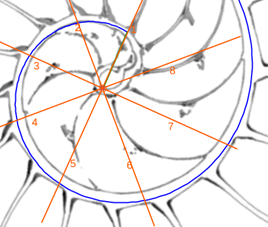
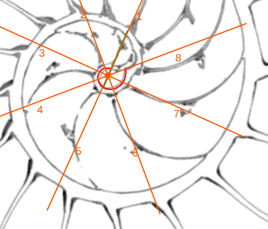
- Con el control tipo botón “centro y radios de paso ” (ver figura 27), disponemos de un conjunto de radios distribuidos con esa amplitud angular constante y con el ratón es posible desplazarlos a voluntad sin más que mover el centro o punto de intersección de ellos y, también, se pueden girar con el pulsador situado a la derecha de ese botón. Con esta herramienta podemos tratar de localizar, si existe, el punto de vista desde el que los apoyos de los septos sobre el labio dorsal y ventral se observen con la esperada o deseada distribución uniforme de paso. Esta búsqueda viene impuesta por el hecho que desde el polo de la espiral ventral no se cumple esa relación (ver figura 28) ni para los apoyos dorsales, ni para los ventrales; y si consideramos esos radios con centro el polo del labio dorsal los apoyos dorsales (ver figura 29) puede considerarse quedan próximos a esa distribución, pero no ocurre así con los ventrales. Le invitamos a realizar sus pruebas e indagaciones usando dicha escena interactiva 4.
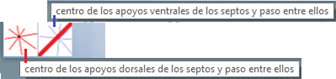
- En las pruebas que he realizado concluyo que hay dos posiciones que modelan adecuadamente esos apoyos septales con distribución uniforme de paso . Ambas las tenemos accesibles respectivamente con el botón etiquetado como “centro de los apoyos dorsales de los septos y pasos entre ellos” y con el botón análogo para “los apoyos ventrales” (ver figura 30). Ambos modelos están reflejados en las figuras 31 y 32. En ellas puede verse que:
- El polo del labio dorsal (punto rojo en la figura 31) y el centro considerado para los radios (punto rojo con borde negro en esa figura) no coinciden, si bien ambos están ubicados en el eje polar (recta dibujada en color verde que une el polo dorsal y el ventral ) y el radio correspondiente al primer septo está alineado con él (recordemos que el primer septo intersecaba al labio dorsal en la constricción y el eje polar también). Las coordenadas determinadas para ese centro, que denominaremos a partir de ahora como centro dorsal, son:
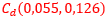 .
. - El polo de la pared ventral (punto azul en la figura 32) y el centro considerado para los radios (punto azul con borde negro en esa figura) claramente no coinciden, pero también ambos están ubicados en el eje polar. El radio correspondiente al apoyo ventral del primer septo está desviado un ángulo aproximado de respecto a dicho eje polar. Las coordenadas de ese centro, que denominaremos centro ventral, son:
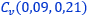 .
.
- El polo del labio dorsal (punto rojo en la figura 31) y el centro considerado para los radios (punto rojo con borde negro en esa figura) no coinciden, si bien ambos están ubicados en el eje polar (recta dibujada en color verde que une el polo dorsal y el ventral ) y el radio correspondiente al primer septo está alineado con él (recordemos que el primer septo intersecaba al labio dorsal en la constricción y el eje polar también). Las coordenadas determinadas para ese centro, que denominaremos a partir de ahora como centro dorsal, son:
 |
|
|
Fig. 27. Botones para situar y girar un haz de radios secantes de paso |
|
 |
 |
|
Fig. 28. Radios de paso |
Fig. 29. Radios de paso |
 |
|
|
Fig. 30. Botones para mostrar los haces de radios de paso |
|
 |
 |
|
Fig. 31. Radios de paso |
Fig. 32. Radios de paso |
Adicionalmente, si mostramos conjuntamente el polo dorsal 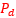 y el ventral
y el ventral 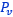 junto a los respectivos centros dorsal y ventral (ver la figura 33), se observa que la mediatriz del segmento
junto a los respectivos centros dorsal y ventral (ver la figura 33), se observa que la mediatriz del segmento 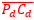 coincide con la del segmento
coincide con la del segmento 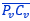 , es decir, que hay un punto C que es centro de simetría de esas dos parejas de puntos.
, es decir, que hay un punto C que es centro de simetría de esas dos parejas de puntos.
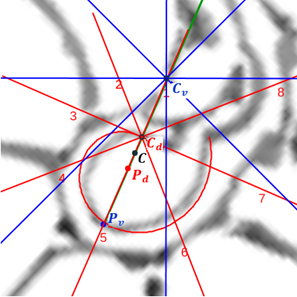
Fig. 33. Alineación de los polos ventral y dorsal con los centros dorsal y ventral. Ubicación simétrica.
Ahora sí, podemos visualizar por qué las cámaras septales aparentan a primera vista una amplitud no uniforme y también por qué las superficies de las secciones de las cámaras se muestran tan diferentes. De nuevo, ya lo detectamos con el sifúnculo, la no coincidencia entre el polo dorsal y ventral que ahora se refleja (reflejo en sentido estricto dada la simetría detectada) en la existencia de dos centros no coincidentes da explicación matemática a esa diferente distribución de cámaras septales en este primer verticilo.
Sobre la superficie de las secciones de las cámaras septales en el primer verticilo
Para analizar el crecimiento de las cámaras y así tratar de detectar los aspectos alométricos[5] que aquí acontecen, hemos construido una herramienta auxiliar que nos permite calcular el área de cada una de las secciones de esas cámaras (ver el espacio de la derecha en la escena interactiva 4 y ver las figuras 34 y 35). Mediante la concanetación de triángulos se puede ajustar la superficie deseada y se facilita de manera automática su área a la escala real del Nautilus.
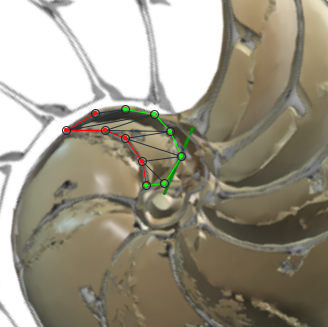 |
|
| Fig. 34. Botón de acceso al medidor de superficies. | Fig. 35. Midiendo el área de una sección de una cámara. |
En la tabla I se refleja la medición realizada con dicha escena interactiva. La columna "Cámara" se corresponde con la numeración de las mismas, "Nautilus 1" recoge las áreas de las secciones del Nautilus Dundee y "Nautilus 2" las del otro ejemplar. El primero cuenta con treinta y tres cámaras y el segundo treinta y dos (en los gráficos comparativos no consideraremos la cámara 33). Las columnas "√pc1" y "√pc2" reflejan respectivamente la raíz cuadrada de las proporciones existentes entre las cámaras pc = An+1/An, 1 ≤ n ≤ 31 para cada uno de los ejemplares.
Dado que el crecimiento angular teórico de la pared ventral en el segundo y tercer verticilo es , y que en base a ello (para comprobarlo puede usarse (14)) se verifican la proporciones:
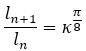 (15)
(15)
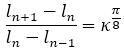 (16)
(16)
donde ln es la longitud de la pared ventral hasta la cámara n y, por tanto, ln+1 - ln es el incremento ventral que ocurre en la cámara n+1, es de esperar que la proporción entre las áreas de las cámaras sea:
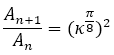 (17)
(17)
Y en el primer verticilo también sería de esperar la obtención de relaciones análogas en base al crecimiento angular de , pero quizás con matices diferenciados pues centros angulares y polos en este verticilo son distintos y ya estamos acostumbrándonos a la diferenciada ontogenia de esta etapa.
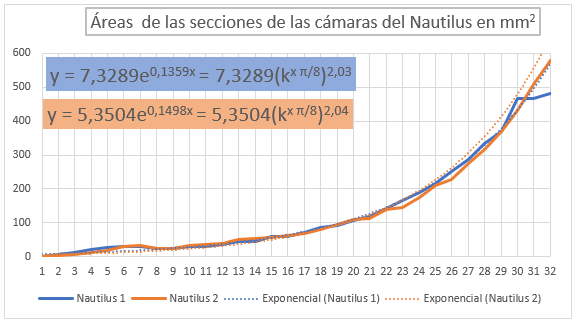
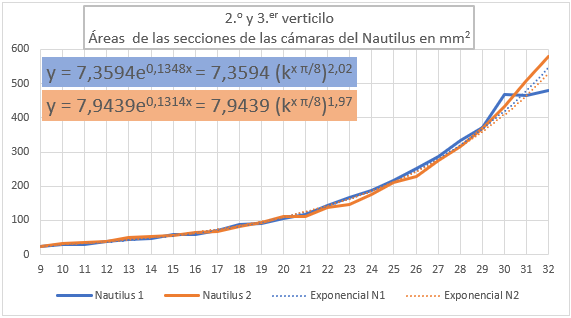
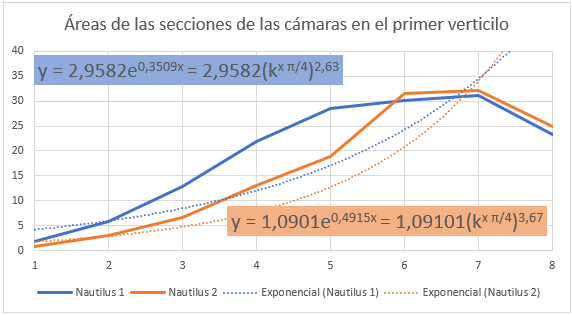
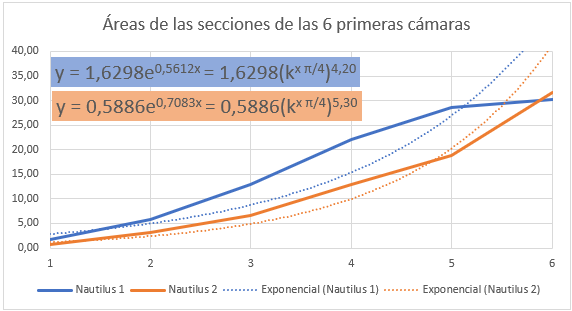
En las figuras 36 y 37 se han representado las áreas de las secciones de las cámaras mediante una poligonal y a su vez se ha determinado la línea de tendencia mediante un ajuste exponencial. En la primera de esas dos gráficas se han considerado todas las cámaras y en la segunda las correspondientes al segundo y tercer verticilo, como hemos señalado el objetivo es detectar posibles variaciones entre la primera etapa de crecimiento y las posteriores. En el primer caso la proporción entre las áreas de cada dos cámaras consecutivas es un poquito superior al valor teórico esperado indicado en (17), pues el exponente en ambos casos es algo superior a dos. Esto está provocado por el crecimiento diferenciado que acontece en el primer verticilo, ya que si consideramos la proporción citada sólo en las cámaras del segundo y tercero (figura 37) entonces sí se está en el entorno cuadrático que caracteriza a ese valor teórico.
En la figura 38 se refleja ese comportamiento diferenciado del primer verticilo y en especial la disminución que acontece en la octava cámara. Si realizamos un ajuste exponencial en este caso el valor esperado sería:
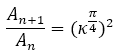 (18)
(18)
al ser el paso angular , pero podemos observar en el ajuste que el valor el superior, e incluso si descartamos la octava cámara (figura 39) el exponente es aún mayor, superior a cuatro.
|
|
Fig. 36. Áreas de las cámaras del Nautilus en los tres verticilos. Ajuste exponencial. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Fig. 37. Áreas de las cámaras del Nautilus en el segundo y tercer verticilo.Ajuste exponencial. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Fig. 38. Áreas de las cámaras en el primer verticilo.Ajuste exponencial. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Fig. 39. Áreas de las seis primeras cámaras del primer verticilo. Ajuste exponencial. |
Pero en este primer verticilo el ajuste más certero sería el logarítmico (ver figura 39) ya que inicialmente en este verticilo el Nautilus ha de conseguir la flotabilidad mediante un crecimiento rápido, es decir, con cámaras amplias y conseguida ésta cierta estabilización, retomando el crecimiento a partir de la cámara novena, pero en este caso de tipo exponencial.
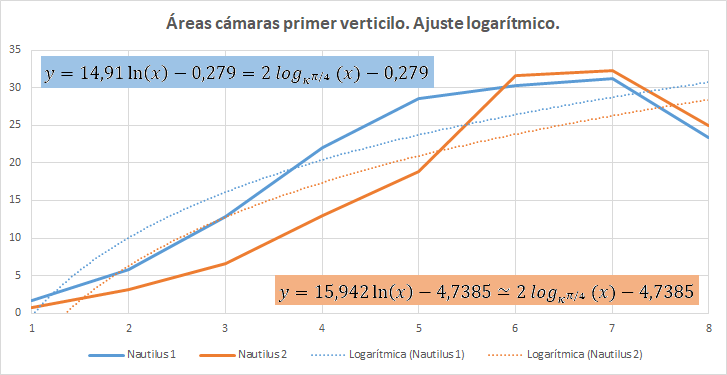
Fig. 39. Áreas de las seis primeras cámaras del primer verticilo
En el ajuste logarítmico del primer verticilo la relación obtenida entre las cámaras es:
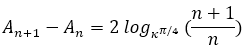 (19)
(19)
Para poder tratar de comprender y explicar lo que acontece será necesario adentrarnos en otra tarea que está intrísecamente relacionada con ésta ya que cada cámara queda determinadas por dos septos y un arco de la pared ventral y otro de la dorsal. Así pues, como ya adelantamos, necesitamos profundizar en los septos, en particular en su amplitud y en especial en el primer verticilo, y también cómo se intersecan estos con la paredes de la concha. Sobre la forma de los mismos sabemos que son arcos de espirales cordobesas. Así pues, ¡adentrémonos!... pero será en un artículo adicional, pues éste ya alcanzó una extensión suficiente.
Bibliografía
Galo J.R., Cabezudo A. y Fernández I.(2016 a) : Sobre la forma y crecimiento cordobés del Nautilus Pompilius. Epsilon, 2016, Vol. 33 (3), nº 94.
González-Restrepo, F. (2019): Cortes del Nautilus a partir de la digitalización 3D del museo Dundee. Red Descartes Colombia.
Greenwald L., Ward P.D. (2010) Buoyancy in Nautilus. In: Saunders W.B., Landman N.H. (eds) Nautilus. Topics in Geobiology, vol 6. Springer, Dordrecht.
https://doi.org/10.1007/978-90-481-3299-7_34
Landman, N. H., Arnold, J. M. and Mutvei, H. 1989: Description of the embryonic shell of Nautilus belauensis. American Museum Novitates, no. 2960, p. 1–16.
[1] Al ser un crecimiento gnomónico, la longitud del apoyo dorsal necesaria para cada cámara va incrementándose, pero aquí lo que buscamos es poner de manifiesto el escaso espacio existente y, consecuentemente, este razonamiento basado en el cálculo del espacio medio disponible es suficiente para alcanzar este objetivo.
[2] Desde el punto de vista causal podríamos apuntar que el hecho de que el espacio disponible sea pequeño es precisamente lo que conduce a ese menor número de cámaras septales. El Nautilus, para construir un nuevo septo ha de desplazarse hacia adelante en su cámara habitacional, necesita un espacio interseptal mínimo, y ese espacio es el que adquiere para cumplir su objetivo natural de crecer y, si es posible, hacerlo gnomónicamente para mantener sus proporciones, su forma. Su preocupación o necesidad es la de avanzar para crecer, ocupando un nuevo espacio, y deja la contabilidad para otros o más precisamente para nosotros.
[3] González-Restrepo (2019) nos aportará próximamente el volumen de cada cámara a partir de los datos de la digitalización aportada para el Nautilus del museo de D’Arcy Thompson.
[4] Al igual que acontecía con el eje del fragmacono, el retardo diferente entre pared dorsal y ventral, 4π vs. 2π, vuelve a aparecer ahora ligado a las cámaras septales.
[5] La alometría son los cambios de dimensión relativa de unas partes corporales en relación a los cambios que acontecen en el tamaño total. Y en particular, la alometría en el crecimiento detecta qué partes de un cuerpo o ente tienen un comportamiento diferenciado.

Este obra está bajo una licencia de Creative Commons Reconocimiento-NoComercial-SinObraDerivada 4.0 Internacional

Para este nuevo curso académico 2021 – 2022 el 'Proyecto Descartes' ofrece una amplia cantidad de objetos informáticos, dinámicos e interactivos de ayuda a la enseñanza y al aprendizaje de los contenidos curriculares de diferentes asignaturas de Bachillerato, fundamentalmente de Matemáticas.
Una de las características de la #REDDescartes es la continua atención a todo lo relacionado con las nuevas tecnologías y la educación de ahí que esté analizando ininterrumpidamente nuevas posibilidades didácticas y metodológicas en este ámbito. La mayor parte de los materiales didácticos de #REDDescartes llevan implícitas técnicas de autoformación y autoevaluación.
El interés actual del Proyecto Descartes queda perfectamente resumido en los siguientes vínculos: enlace 1 y enlace 2.

El Proyecto Descartes ha creado recursos interactivos para PC, tablet y smartphone en las áreas de: Matemáticas, Lengua, Ciencias Naturales, Física y Química, Biología, Inglés, Francés, Artes Plásticas, Tecnología, Geografía, etc. Ver:
https://proyectodescartes.org/indexweb.php
En la página enlazada anteriormente se muestran todos los recursos disponibles hasta el momento del acceso. Se observa que los relativos a Bachillerato se encuentran en los subproyectos:
|
|
 Misceláneas. Matemáticas Secundaria, Bachillerato y Universidad |
 Libros interactivos de Descartes Libros interactivos de Descartes |
 Proyecto Un_100: Matemáticas y Física. Bachillerato y Universidad |
 Problemas: Física y Química. Secundaria, Bachillerato y Universidad |
 Proyecto Prometeo. Matemáticas y Física |
 Juegos: Todos los niveles y asignaturas |
 Ingeniería y Tecnología |
 Estudio Europeo de comunicación lingüística |
| Unidades de Geografía |

En algunos subproyectos no se menciona explicitamente que contengan material para bachillerato pero se han añadido por considerar que parte de los recursos de dichos subproyectos son aplicables a la etapa.
Hemos hecho notar que la disponibilidad de materiales está actualizada hasta el momento del acceso debido a que, en la actualidad, el proceso de creación de nuevos recursos está experimentando un crecimiento extraordinario tanto en cantidad como en calidad y diversidad, lo que hace aconsejable revisar con cierta frecuencia los recursos disponibles. En la actualidad se está adaptando el 'Proyecto Prometeo', abundante en contenidos y el proyecto iCartesiLibri versatil y de profundo interés, con lo que el aporte de nuevo material es contínuo.
Si bien los libros del subproyecto iCartesiLibri son creaciones dinámicas e interactivas pensadas para usarse tal cual están, al igual que los recursos del resto de los subproyectos, es de señalar la posibilidad de adaptar fácilmente los contenidos de dichos recursos a las necesidades propias o a las características del aula, especialmente las Misceláneas que con un mínimo de cambios pueden convertirse en herramientas propias que satisfagan las necesidades didácticas inmediatas. Todo lo que necesitamos para realizar los cambios en los recursos es el 'Editor Descartes' herramienta (editor) intuitiva y de fácil uso.
Una vez que se ha visto la estructura de un recurso y se ha hecho uso de él se intuye la facilidad para la creación de uno parecido para lo cual disponemos de tres libros interactivos en el subproyecto iCartesiLibri, 'Formación en DescartesJS', que detallan todas las funcionalidades y procedimientos del editor Descartes y también está disponible una documentación exhaustiva en línea.

Cada subproyecto y cada recurso del mismo disponen de la documentación necesaria para que fácilmente identifiquemos sus objetivos y las indicaciones necesarias para su uso.
Estamos hablando de los recursos digitales que podemos encontrar en el Proyecto Descartes y de planificación de un curso escolar en estado de pandemia donde es importante disponer de un método de trabajo que pueda facilitar la formación de alumnos cuando se dificulta o incluso se suprime la enseñanza presencial. Recomiendo el informe de la experiencia de nuestro compañero José Antonio Salgueiro, hábil usuario de las técnicas de comunicación, cuando en el curso pasado se vio obligado a cerrar su aula e interrumpir la enseñanza presencial con un grupo de alumnos. El resultado de su experiencia bien puede servir de inspiración al docente o al menos tenerlo en cuenta al hacer su propia planificación.
En su artículo titulado "Evaluación, durante el estado de alarma, de la práctica docente a distancia con un modelo sin videoconferencia" nos dice
"En primer lugar, dejaremos constancia de que este artículo no se fundamenta en ningún trabajo de investigación educativa ni en una experimentación didáctica previamente planificada y diseñada con unos objetivos concretos, sino que pretende difundir entre el claustro virtual una experiencia surgida de la improvisación y compartir los recursos generados con la práctica docente a distancia y sin videoconferencia en la materia de Matemáticas-II del Bachillerato de Ciencias con el alumnado del IES Bajo Guadalquivir de Lebrija, por si fuera de utilidad para otros compañeros y compañeras."
El autor ha publicado, recientemente, otros artículos y experiencias de indudable interés para todo docente interesado en la metodología en tiempos "difíciles".
Ildefonso Fernández Trujillo. 2021

La esencia del Nautilus es ostensiblemente cordobesa, pero al analizar su ontogenia biológica se detecta un comportamiento diferente en cada uno de los tres verticilos que pueden contabilizarse en la concha de todo animal adulto. Esta diferencia es especialmente manifiesta y significativa en el primero de ellos. En este artículo, nos ponemos nuestras gafas matemáticas para tratar de localizar la causa y así poder mejorar el modelo matemático uniforme que previamente habíamos compartido. Aquí, nos adentramos plenamente en la ontogenia matemática del Nautilus. Toda espera queda satisfecha para quienes son pacientes y están interesados.
En los artículos previos (Ontogenia I y Ontogenia II) llegamos a detectar la necesidad de una revisión del modelo cordobés uniforme del Nautilus, en especial en el primer verticilo. Y para lograr modelar la ontogenia matemática citada, tendremos que apoyarnos en la ontogenia biológica detectando en ésta los diferentes detalles y particularidades que determinan las características esenciales en cada momento y etapa, y tendremos que concretar cada una de ellas para su adecuada traslación al modelo matemático final. Esto es lo que abordaremos a continuación.
Observando la ontogenia biológica del Nautilus con gafas matemáticas
Concha embrionaria
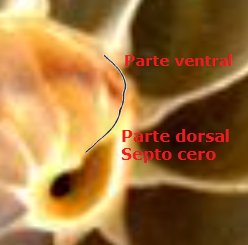
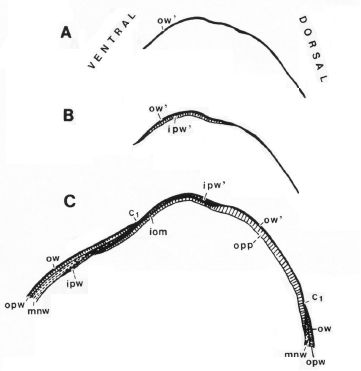
La concha embrionaria inicial, aquella que no tiene ninguna cámara y septo, y de ahí que se le denomine preseptal, tiene aspecto de lapa (Figuras 7 y 10). Su perfil (imagen A de la Fig. 11 y Fig. 15) queda modelado por dos curvas: una que se corresponde con la parte ventral, aquella que tendrá continuidad en la pared ventral de la concha, y otra dorsal que podría considerarse como el septo inicial o septo cero (Mutvei & Doguzhaeva, 1997). Esta característica primigenia se traslada y mantiene permanente en todas la etapas vitales, es un invariante, pues como conocemos se verifica que todo punto interior a la concha o sobre ella se obtiene como la intersección de dos curvas espirales, una longitudinal similar a la ventral y otra transversal similar a la septal.
Curvatura ventral
La constricción de la cicatriz, coincidente con la intersección de la concha embrionaria con el primer septo, es la que marca el inicio del patrón de crecimiento correspondiente a la sección de la concha ventral. A partir de la imagen correspondiente a la fase del primer septo (ver Fig. 16) podemos observar claramente la curvatura de la parte ventral y verificar cómo ésta se ajusta a una espiral cordobesa, ajuste que permanece durante toda la vida del animal (Fig. 17). Consecuentemente, ésta es una característica intrínseca y permanente, un segundo invariante, en toda la ontogenia del Nautilus desde el periodo embrionario septal a la madurez.
 |
 |
 |
| Fig. 15. Perfil de la concha embrionaria o preseptal | Fig. 16. Ajuste de la pared ventral en la fase del primer septo. C1: Constricción |
Fig. 17. El perfil ventral es cordobés en toda la etapa vitall |
Tangencialidad de los septos con la pared ventral de la concha
En las imágenes de las figuras 12, 13 y 14 puede intuirse que las paredes septales confluyen con la pared ventral de la concha tangencialmente, lo cual a priori parece lógico que acontezca para que el cuerpo del animal pueda asentarse de manera uniforme en la zona de unión de ambas paredes. Eso se visualiza claramente a nivel microscópico (Fig. 18), donde en la parte apical (parte izquierda en esa imagen) también puede observarse un cemento de unión entre ambas paredes que a nivel macroscópico difumina esta tangencialidad y puede inducir a interpretar inadecuadamente la forma en que ambas capas confluyen. Esta tangencialidad se da en toda la ontogenia del Nautilus, tercer invariante, y necesitamos destacarla y fijarla porque es básica para el modelo matemático buscado.
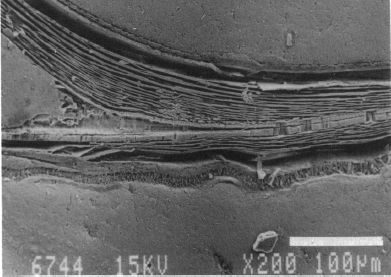
Fig. 18. Detalle de la unión del segundo septo y la parte ventral de la concha en la fase de dos cámaras. Ampliación de la imagen de la Fig. 13. Entre ambas paredes en la parte apical se observa un cemento de unión. (Procedente de Landmam et al.1989).
Incidencia de los septos con la pared dorsal de la concha
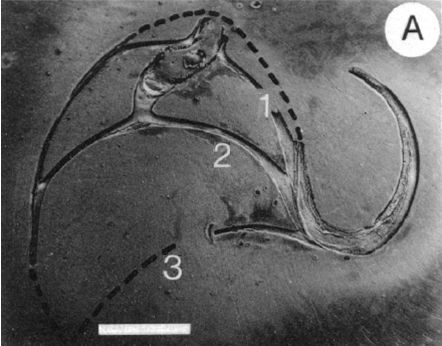
La modelación de la incidencia de los septos con la pared dorsal requiere un análisis detallado ya que presenta variaciones destacables en cada verticilo y, por tanto, habrá que localizar la causa de las mismas. Así pues, observando la unión de los dos primeros septos con dicha pared dorsal (Fig. 19) podemos constatar que aquí no acontece, no se reproduce, la indicada tangencialidad ventral y se detecta cemento de unión en la parte apical (en esta imagen ubicada a la derecha) y en la adoral (izquierda). Matemáticamente, si partimos de que la forma del septo sigue una determinada curva prefijada (en el modelo global uniforme indicamos, mostramos y asumimos que es un arco de espiral) y considerando la tangencialidad indicada de éste con la pared ventral, entonces se concluye que la incidencia que acontece con la pared dorsal viene dada, está preestablecida, es forzada y tiene que asentarse según el ángulo que corresponda en cada momento, y quizás ello es lo que constructivamente obliga a que la unión se tenga que reforzar para obtener una consistencia adecuada (de ahí el cemento indicado). Algebraicamente, detectada la forma de la de la pared dorsal, la determinanción del ángulo y punto de incidencia quedan reducidos a intersecar la ecuación del septo con la de la pared dorsal, y eso es lo que haremos en su momento y en cada una de las fases del crecimiento, las cuales hemos asociado a cada verticilo.

Fig. 19. Detalle de la unión de los dos primeros septos (1 y 2) y la parte dorsal de la concha en la fase de dos cámaras. Ampliación de la imagen de la Fig. 13. Entre ambas paredes se observa un cemento de unión. (Procedente de Landmam et al.1989).
Adicionalmente, dado que la parte dorsal de la concha es diferente en el primer verticilo (lo analizaremos posteriormente en un apartado específico), la amplitud del arco del septo parece ser variable y ,si es así, tendrá que acortarse o alargarse hasta encontrar el punto de apoyo necesario para el cierre de cada cámara. Acorde con ello, el ángulo de incidencia observado parece ser diferente en los septos del primer verticilo (observar detenidamente la Fig. 20, imagen A). En esa misma figura se observa que en el segundo verticilo, en el área dorsal media (mda), son ostensibles las depresiones septales dorsales (dsd), ver detalle de su forma en las imágenes B, C y D, y éstas se vuelven menos notables en el tercer verticilo. La ontogenia biológica parece mostrarse esquiva con la matemática o simplemente gusta de marcarnos retos adicionales sobre los que dilucidar.
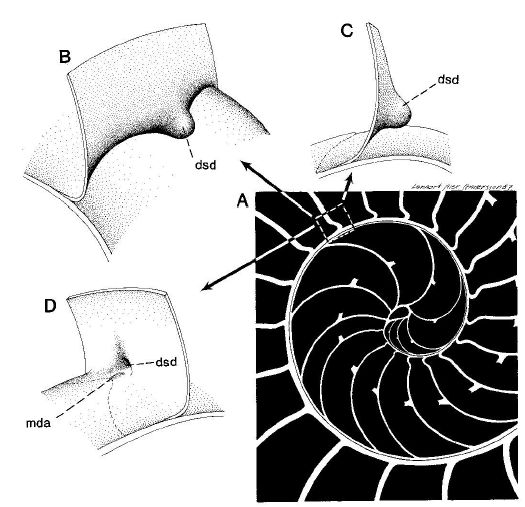
Fig. 20. Sección de la concha del Nautilus. Diferentes vistas de la depresión septal dorsal (dsd) en el área dorsal media (mda). (Procedente de Mutvei & Doguzhaeva, L. 1997).
Labio dorsal y ombligo
Como indicamos con antelación (ver la figura 10) es en la constricción dorsal, que es también el punto de incidencia del primer septo, donde el perfil de la concha en su parte dorsal cambia su concavidad y adopta la misma que la pared ventral, si no fuera así, la cavidad se cerraría. Y al confluir la pared dorsal de este primer verticilo con la concha embrionaria preseptal es como se forma el ombligo.
A priori, no es fácil poder modelar con certeza la forma que adopta este labio dorsal dado que su recorrido es inferior a una vuelta y, consecuentemente, las referencias sobre las que apoyarse puede que no sean suficientes. Lo que sí puede observarse es un posible crecimiento en espiral, es esperable que sea así ¿verdad?, y además que ésta sea una espiral geométrica para que el crecimiento sea gnomónico; por tanto, planteamos y establecemos esta hipótesis. Adicionalmente, por uniformidad, consideraremos que el tipo de crecimiento sea también cordobés ya que la pared dorsal en el segundo y tercer verticilo tiene esa forma (las paredes dorsales en estos verticilos eran previamente, en la vuelta anterior, sendas paredes ventrales ) y es lógico que ésta se mantenga en el primero o mejor dicho que ésa sea la tendencia original que continúa posteriormente. Es esperable y lógico que la curvatura del dorso del animal se mantenga en toda su existencia. Bajo esa consideración, lo que sí es obvio (observar la Fig. 16) que:
- La espiral que modela la pared ventral no modela al labio dorsal. Observad como la parte correspondiente a esta espiral que se ha dibujado en la imagen mediante puntos no se ajusta a dicho labio.
- El polo de la espiral del labio no puede coincidir con el polo de la espiral de la pared ventral ya que el primero ha de ubicarse en el interior del ombligo y el segundo según lo ya modelado se sitúa sobre su borde (ver Fig. 21). Estamos hablando de una distancia entre esos dos polos que estará en el entorno de un milímetro y que puede parecer una diferencia no significativa, sin embargo, como veremos, ésta es realmente la clave diferenciadora y la causa de las variaciones que se observan en el crecimiento durante el primer verticilo respecto al de los restantes. Sí, ¡así es de simple!. A nivel global, a la escala de la concha de un ejemplar maduro, no hay una diferencia visual entre ambas (ver en la esquina superior derecha en la figura 21 que la espiral ventral, en color azul, y la dorsal en rojo se "solapan"), pero a nivel local se contempla un comportamiento muy diferenciado (ver la parte central en esa figura 21), ¡esas son las maravillas de la espiral logarítmica o geométrica o "Spira mirabilis" como la bautizó Jacob Bernoulli.
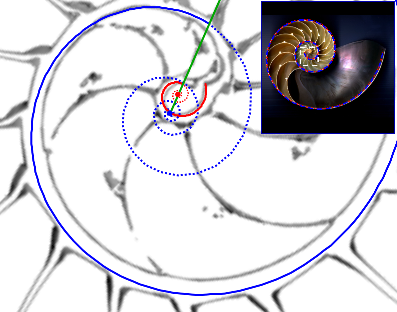
Fig. 21. Modelización de la concha en el primer verticilo. Espirales cordobesas con diferente polo. En rojo el labio dorsal y en azul la pared ventral. Diferenciado a nivel microscópico (imagen central) y "coincidente" a nivel macroscópico (esquina superior derecha).
¡Vaya! Ya lo había adelantado literariamente antes, pero ahora ¡se me hizo irresistible mostrarles gráficamente, en esta figura 21, la causa o germen que permitirá comprender mejor al Nautilus! Bien sabe quien se dedica a investigar que no puede ocultarse la satisfacción cuando en el interior de nuestra mente surge la inspiración ¡Ajá! que sublimó Martin Gardner y la que llevó a Arquimedes a correr desnudo, insconcientemente, a la par que gritaba ¡Eureka! a sus paisanos de Siracusa. Y salvando las distancias y comparaciones, estaba deseoso de que lo vieran. Pero esa inspiración inicial, que esporádicamente regalan las musas generalmente sólo a quien está trabajando, no es más que la piedra angular que asienta y modula la futura estructura global, pero habrá que seguir construyendo sobre ella en base a nuevas inspiraciones y deducciones lógicas. Así pues, hemos de asentar esa piedra base y aplicar todos los detalles detectados, analizados y descritos en la particular revisión que hemos realizado de la ontogenia biologica del Nautilus, para así llegar con paso firme a nuestro objetivo que es poder comunicarles lo que sotto voce nos cuenta el Nautilus acerca de su ser y crecer, y poder hacerle de traductor al lenguaje que muchos catalogan como el criptográfico o secreto de la Naturaleza: el matemático. Ahora sí, llegó el momento esperado, o al menos anunciado, de formular detalladamente la Ontogenia matemática del Nautilus. Gracias por ser pacientes, considero que se verán gratificados por haber llegado aquí y por continuar conmigo.
Ontogenia matemática del primer verticilo del Nautilus
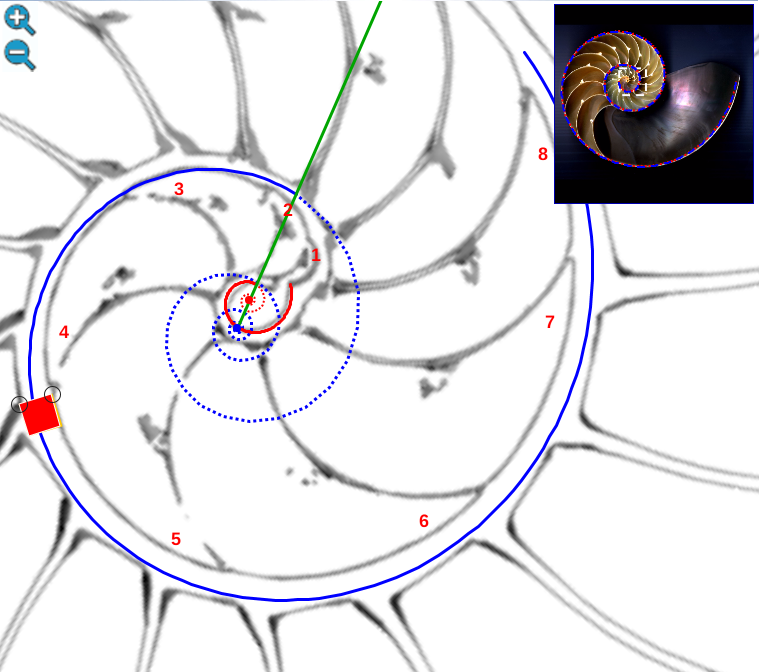
Para realizar el análisis matemático en este primer verticilo utilizaremos el siguiente recurso interactivo (escena interactiva 2):
Escena interactiva 2. Concha y sifúnculo del Nautilus en el primer verticilo.
Puede interactuar libremente con ella. Si posiciona el ratón sobre los botones tendrá una breve información o consulte las instrucciones.
Modelando el labio dorsal
Para mostrar la modelización del labio dorsal y el primer verticilo hemos elegido la concha digitalizada por el Museo de D’Arcy Thompson (Fig. 22) en la Universidad de Dundee y, en particular, los datos tridimensionales aportados por dicho museo correspondientes a esa digitalización. A partir de ellos, González-Restrepo (2019) obtuvo la sección sagital y las parasagitales (Fig. 23), así como las radiales. Realizamos esta elección porque en esta fuente gráfica el corte con los septos del primer verticilo y con los restos del canal sifuncular es más visible que los que pueden observarse en la concha que hemos utilizado, hasta ahora, como soporte de referencia para mostrar el modelo uniforme y las propiedades observadas. Aquí se puede ver que los septos del primer verticilo están incompletos y consecuentemente tampoco se visualiza el rastro del canal del sifúnculo, por tanto, no es posible modelar lo que no es visible. Esta es la razón de cambiar la referencia gráfica que utilizamos como muestra.
| Modelo tridimensional de la sección de la concha de un Nautilus pompilius. |  |
|
Fig. 22. Nautilus Shell (sectioned) by University of Dundee Museum Collections on Sketchfab |
Fig. 23. Cortes por planos paralelos al plano medio obtenidos por González-Restrepo (2019) a partir del modelo 3D del Museo Dundee. |
Partamos de que la espiral logarítmica cordobesa[1] que modela la pared ventral (representada en color azul en las imágenes y escenas) viene dada en coordenadas polares por:
![]() (1)
(1)
o en coordenadas cartesianas en función del parámetro θ :
 (2)
(2)
es decir, que ubicamos su polo en el origen de coordenadas. Adicionalmente denotaremos el ángulo inicial de la espiral como θi, y como θf el ángulo final correspondiente a la boca de la concha.
Con la notación anterior, entonces el polo de la espiral cordobesa que aproxima al labio dorsal se situaría en D(0.035, 0.080) y su ecuación paramétrica sería:
 (3)
(3)
Donde el coeficiente ![]() , verificaría que
, verificaría que ![]() .
.
De los valores posibles el que teóricamente podría ser más certero se correspondería con:
![]() (4)
(4)
pues matemáticamente representaría que el labio dorsal tiene un retraso de dos vueltas (un ángulo de retardo de 4π) respecto a la pared ventral inicial. Sería de esperar que el retraso fuera de 2π, pues ésta es la relación que acontece en los siguientes verticilos cuando la que antes era pared ventral pasa a ser la nueva pared dorsal en la siguiente vuelta (de ahí lo de 2π), pero lo esperado no es siempre lo encontrado.
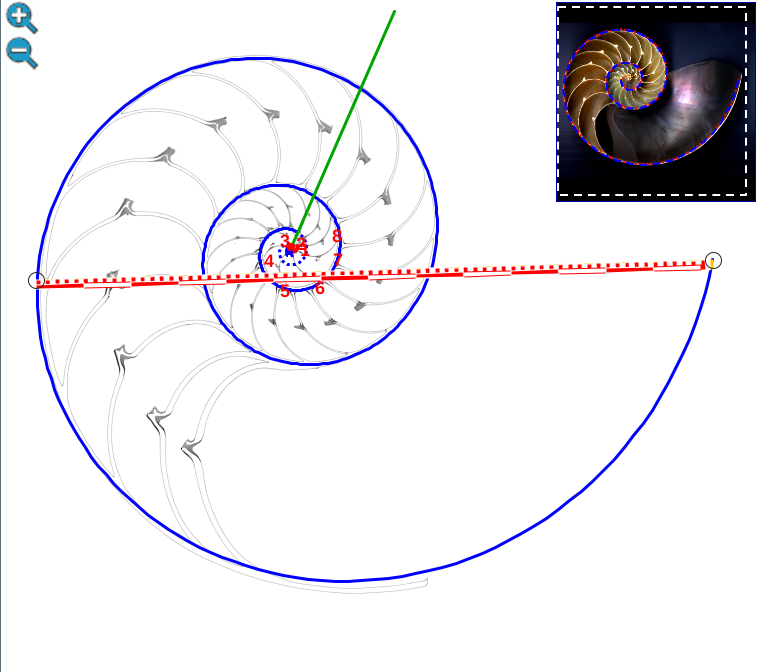
Geométricamente, si el labio dorsal en el primer verticilo tuviera sólo el retardo de 2π se generaría un ombligo de mayor tamaño al que se observa en la concha del Nautilus. Esto representaría al animal un esfuerzo constructivo mayor, pues tendría que cubrir una superficie más amplia (en este caso más longitud dado que estamos trabajando con la sección) ya que este labio dorsal bordearía a la concha embrionaria. En la escena interactiva 3, espacio izquierdo, puede verse cuál sería la forma del labio dorsal, ombligo y concha en el supuesto de un retardo de 2π y en el espacio derecho el caso de retardo 4π. Entre las dos alternativas la correspondiente a 4π parece la más eficiente. Podrían plantearse retardos que no tuvieran que ser múltiplos de 2π, pero de partida ello no se observa como biológicamente adecuado, pues estaría considerándose que la parte dorsal del cuerpo del animal tendría un crecimiento descompensado, al menos en algún instante o periodo temporal, respecto a la ventral. Pudiera acontecer, pero el deslizamiento que realiza a lo largo del fragmacono al crecer, inspira más una situación más regular. Si en el periodo evolutivo acontecido en la larga historia de este animal han ocurrido retardos de otro tipo, al menos parece que la selección natural ha conducido al retardo de 4π que hemos detectado y que consideramos en nuestro modelo del primer verticilo.
Escena interactiva 3. Modelando el labio dorsal del Nautilus en el primer verticilo.
Puede interactuar libremente con ella. Si posiciona el ratón sobre los botones tendrá una breve información o consulte las instrucciones.
Ambas espirales comparten el mismo eje polar (en color verde en la escena), es decir, el eje de cada una de ellas está incluido en la semirrecta que pasa por el polo ventral y por el polo dorsal. Adicionalmente, esta semirrecta interseca a la concha en la constricción dorsal y la ventral que son los puntos de intersección y tangencia, respectivamente, del primer septo con la pared dorsal y la ventral de dicha concha. Esto cuadra con la ontogenia biológica descrita antes y matemáticamente aporta también una pauta común de crecimiento al compartir ambas espirales el mismo parámetro angular. El ángulo inicial común para ambas espirales indicamos que lo denotaríamos como θi y entonces el recorrido total de la espiral dorsal termina en ![]() .
.
La pequeña variación existente entre los polos de ambas espirales (aproximadamente un milímetro) conduce a que el labio dorsal no finalice su recorrido siendo tangente a la concha embrionaria preseptal, sino que este labio incide e interseca a dicha concha primigenia cuando se cierra el primer verticilo y se genera el ombligo. Consecuentemente, el enlace con la parte ventral inicial no es suave, sino que se produce a través del séptimo y octavo septo. Son estos últimos septos del primer verticilo los que al apoyarse en la concha primigenia se adaptan para dar continuidad con la pared ventral inicial. El hecho de que no se produzca esa tangencialidad entre la pared ventral inicial y la finalización del labio dorsal refuerza y muestra la necesidad de considerar la existencia de dos polos diferenciados en el modelo teórico y, también, que no se pueda atribuir esta diferencia a un error en la precisión del ajuste gráfico realizado[2]. Ver detalle del modelo a nivel local en la figura 22 y a nivel global en la figura 23.
 |
 |
|
Fig. 22. Ajuste del labio dorsal (espiral en rojo) y de la pared ventral en el primer verticilo (espiral azul)). La sección gráfica de referencia se debe a González-Restrepo (2019). En rojo la regla de medida que marca 1 mm. |
Fig. 23. Ajuste global de la concha del Museo de D’Arcy Thompson, en base al corte realizado por González-Restrepo (2019). La regla de medida marca 143 mm. |
El canal sifuncular en el primer verticilo
En nuestro primigenio modelo uniforme detectamos que el canal sifuncular actuaba como el eje del fragmacono, es decir, que es equidistante de la pared ventral y dorsal. En este supuesto uniforme ambas paredes describen sendas espirales cordobesas con igual polo, de manera que si la ecuación en coordenadas polares de la primera es ![]() , la de la segunda sería
, la de la segunda sería ![]() (realmente es la misma espiral, pero la segunda con un ángulo de retardo de 2π o coeficiente 0,3431...) y, entonces, el sifúnculo tiene de ecuación
(realmente es la misma espiral, pero la segunda con un ángulo de retardo de 2π o coeficiente 0,3431...) y, entonces, el sifúnculo tiene de ecuación
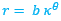 (5)
(5)
donde 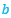 toma el valor medio de los respectivos coeficientes 1 y
toma el valor medio de los respectivos coeficientes 1 y 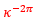 de las espirales anteriores, es decir,
de las espirales anteriores, es decir,
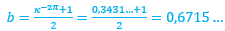 (6)
(6)
lo que se corresponde con una espiral cordobesa con un ángulo de retardo de ![]() . Pero, ya comprobamos que el ajuste anterior es adecuado en el segundo y tercer verticilo, pero no así para el primero. No obstante, ahora sí, estamos en condiciones de determinar cómo queda modelado el sifúnculo en esta primera vuelta.
. Pero, ya comprobamos que el ajuste anterior es adecuado en el segundo y tercer verticilo, pero no así para el primero. No obstante, ahora sí, estamos en condiciones de determinar cómo queda modelado el sifúnculo en esta primera vuelta.
El fragmacono en el primer verticilo está delimitado por dos espirales cordobesas cuyas ecuaciones son (2) y (3), ambas con un ángulo polar común, pero con distinto polo. Si consideramos que el sifúnculo en este verticilo tiene un comportamiento similar a los dos posteriores, entonces éste sería el eje de ese fragmacono y, para determinarlo, procedamos a determinar el lugar geométrico de los puntos del plano que equidistan de ambas y obtendremos que viene determinado por la ecuación:
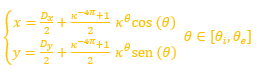 (7)
(7)
que se corresponde con una espiral cordobesa, cuyo polo![]() es el punto medio de segmento determinado por el polo de la pared dorsal
es el punto medio de segmento determinado por el polo de la pared dorsal ![]() y el de la ventral
y el de la ventral ![]() , y su coeficiente es la media aritmética de los coeficientes respectivos[3]
, y su coeficiente es la media aritmética de los coeficientes respectivos[3]
![]() (8)
(8)
Al representar gráficamente esta espiral (ver en la imagen derecha de la fig. 24 la línea discontinua en color naranja o interactuar en la escena interactiva 3) observamos que realmente no modela adecuadamente a dicho sifúnculo en ese verticilo. Sin embargo, si observamos en esa figura 24 la espiral trazada en una línea continua, también de color naranja, ésta sí se ajusta al los restos del canal sifuncular.
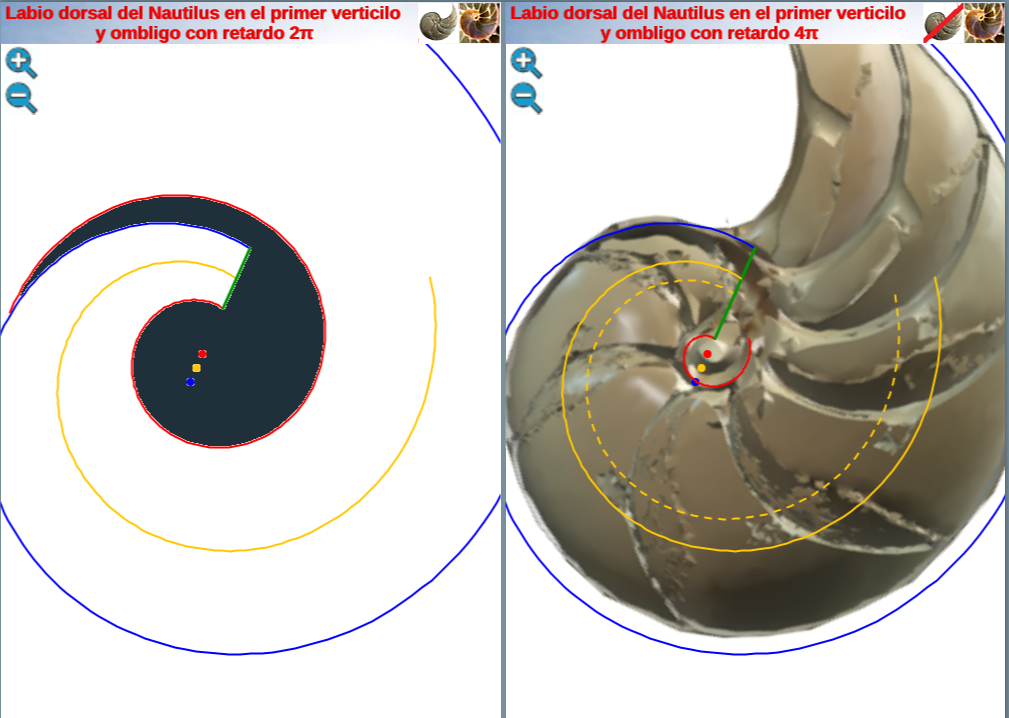
Fig. 24. En la imagen de la izquierda y de la derecha eje del fragmacono en el primer verticilo cuando el labio dorsal tiene un retardo de 2π (espiral continua de color naranja). A la derecha en línea discontinua eje del fragmacono en el primer verticilo cuando el labio dorsal tiene un retardo de 4π. La primera se ajusta al canal sifuncular.
La ecuación de esa espiral y sifúnculo es la siguiente:
 (9)
(9)
donde
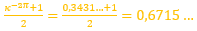 (10)
(10)
y se correspondería con el eje del fragmacono, pero en el caso de que el labio dorsal tuviera sólo un retardo de 2π (ver imagen izquierda en la figura 24 o interactuar en la escena 3). Ese coeficiente o retardo en el sifúnculo es el mismo que acontece en el segundo y tercer verticilo, pero aquí el polo es diferente. En la figura 25 podemos ver una comparativa de la espiral sifuncular aquí determinada (en color naranja) y la del sifúnculo en el modelo uniforme (en turquesa).
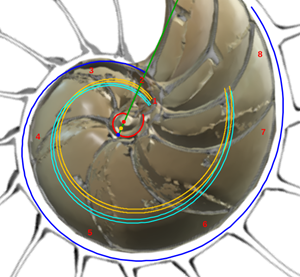
Fig. 25. Comparativa de la espiral sifuncular del primer verticilo (naranja) y la del sifúnculo en el modelo uniforme (turquesa).
Así pues, la causa de que el sifúnculo tenga un comportamiento diferenciado en el primer verticilo se centra en el hecho de que el labio dorsal y la pared ventral tienen diferente polo. La explicación de cuál es la causa de su desviación respecto al eje del fragmacono queda abierta...
Pienso que hemos dado un gran paso en la comprensión y explicación de la ontogenia matemática del Nautilus. En ella, sigue imperando su carácter netamente cordobés, introduciendo bonitos matices matematicos que explican los matices biológicos observados. Pero D. Antonio Machado nos indicó que se hace camino al andar y para ello, después del primer paso dado, han de llegar otros y en nuestro estudio, ahora, tendremos que centrarnos especialmente con las paredes septales y, ¿por qué no?, quizás tratar de localizar cuál es el germen de la esencia cordobesa de este entrañable, al menos para mí, animalito. Así pues, tendré que preparar un nuevo artículo. ¿Será el último de esta serie o no será? El tiempo y la inspiración lo dirá... Les estaré esperando por aquí... ¡pronto!
[1] Para saber por qué esa espiral es una espiral cordobesa, por qué la razón de que la base de esa exponencial sea κ = 1,18558… y su significado, puede consúltarse: Galo et al (2016).
[2] De nuevo, hemos de recordar que el paso de la yocto-yotta realidad al modelo teórico requiere de la consideración de cierta tolerancia, pero que hay que saber distinguirla de la existencia de una diferencia que provoque cambios en el modelo. Si la toma en consideración de esa tolerancia no cambia el modelo podremos aceptarla y admitirla, pero habrá que evitarla cuando se provoque un cambio en él. Así pues, en este caso es obligado considerar que los polos son diferentes. Por el contrario, por ejemplo, en las espirales que hemos considerado para ajustar el perfil del labio dorsal y de la pared ventral, en especial ésta, podría aducirse que necesitarían una mejora en su ajuste a la imagen de la concha en ese primer verticilo, pero realmente no es necesario ya que tal y como se ha considerado es suficientemente adecuado para determinar el modelo global que caracteriza al genus Nautilus. Esto ya lo mostramos en la fig. 21, y lo detallamos ahora, pues al observar el ajuste de la pared ventral (espiral azul) en el primer verticilo y su ajuste global en la toda la concha, se marca de manera evidente que la tolerancia considerada es admisible para determinar el modelo a nivel tanto local como global (el grosor de la pared es inferior a un mm, lo puede medir en la escena y el ejemplar considerado tiene unos 143 mm en su etapa adulta, figuras 22 y 23) . Sin embargo, si modificásemos la posición del polo de la pared dorsal, es decir, ignorásemos la pequeña distancia que existe entre ambos polos obtendríamos variaciones no admisibles a nivel local, es decir, cambiaría el modelo y no se ajustaría a la ontogenia del Nautilus.
[3] En general puede comprobarse que dadas dos espirales logarítmicas con igual base, con el mismo ángulo polar y con diferentes polos; si se halla el lugar geométrico de los puntos de plano obtenidos mediante una combinación lineal de las mismas se obtiene que es una espiral logarítmica del mismo tipo, es decir, de igual base y ángulo polar, y con polo la combinación lineal de los polos y coeficiente la combinación de los coeficientes.
Bibliografía
Galo J.R., Cabezudo A. y Fernández I.(2016 a) : Sobre la forma y crecimiento cordobés del Nautilus Pompilius. Epsilon, 2016, Vol. 33 (3), nº 94.
González-Restrepo, F. (2019): Cortes del Nautilus a partir de la digitalización 3D del museo Dundee. Red Descartes Colombia.
Landman, N. H., Arnold, J. M. and Mutvei, H. 1989: Description of the embryonic shell of Nautilus belauensis. American Museum Novitates, no. 2960, p. 1–16.
Mutvei, H. and Doguzhaeva, L. 1997: Shell ultrastructure and ontogenetic growth in Nautilus pompilius L. (Mollusca: Cephalopoda). Palaeontographica Abteilung A Palaeozoologie–Stratigraphie, vol. 246, p. 33–52.
Este obra está bajo una licencia de Creative Commons Reconocimiento-NoComercial-SinObraDerivada 4.0 Internacional

Título: Cálculo, vol. I. Cálculo diferencial.
Sección: iCartesiLibri
Bloque: Análisis matemático
Unidad: Cálculo diferencial
Nivel/Edad: Universidad (18 años o más)
Idioma: Castellano
Autores: Elena Esperanza Álvarez Sáiz y Juan Guillermo Rivera Berrío
ISBN obra completa: 978-84-18834-18-9
ISBN del volumen: 978-84-18834-19-6
![]() Haz clic aquí para ver una versión en pdf
Haz clic aquí para ver una versión en pdf
![]() Haz clic en la imagen para abrir el recurso
Haz clic en la imagen para abrir el recurso
Puedes encontrar todos los libros interactivos de iCartesiLibri en
https://proyectodescartes.org/iCartesiLibri/index.htm - Ver Créditos
Este material está publicado bajo una licencia:

Creative Commons Reconocimiento-NoComercial-CompartirIgual 4.0 Internacional

Título: Desarrollo humano y social
Sección: iCartesiLibri
Bloque: Ciencias humanas y sociales
Nivel/Edad: Bachillerato y Universidad (16 años o más)
Idioma: Castellano
Autores: Mario Javier Naranjo Otálvaro, Gustavo Adolfo Tobón Pereira, Héctor Eduardo Cardona Carmona, Juan Guillermo Rivera Berrío
ISBN: 978-84-18834-17-2
![]() Haz clic aquí para ver una versión en pdf
Haz clic aquí para ver una versión en pdf
![]() Haz clic en la imagen para abrir el recurso
Haz clic en la imagen para abrir el recurso
Puedes encontrar todos los libros interactivos de iCartesiLibri en
https://proyectodescartes.org/iCartesiLibri/index.htm - Ver Créditos
Este material está publicado bajo una licencia:
Creative Commons Reconocimiento-NoComercial-CompartirIgual 4.0 Internacional

En su primer verticilo, la concha del Nautilus tiene un crecimiento diferenciado del que acontece en el segundo y tercero. Su esencia, implícita y explicitamente, es siempre cordobesa, pero de partida el número de cámaras en ese primer verticilo son ocho y consecuentemente con paso π/4 entre ellas, frente a las diecisés y paso π/8 del segundo, paso que también ocurre en el tercero. ¿Qué acontece en la ontogenia del Nautilus que sea la causa productora de esa distinción? En este segundo artículo (desde aquí puede acceder al primero) nos adentraremos en la biología de este animal, o más precisamente en el rastro calcáreo que deja y que conforma su concha, para tratar de dar respuesta matemática a esta pregunta. Procedamos a iniciar y a adentrarnos en la ontogenia del Nautilus y desde ahí trascender a su ontogenia matemática.
La concha embrionaria y el primer verticilo
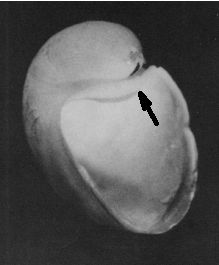
A primera vista la concha embrionaria del Nautilus (Fig. 6), en su periodo pre-cameral, aparenta una forma similar a la concha de una lapa (Fig. 7) (MolluscaBase eds., 2021), pero a diferencia del crecimiento que sigue ésta, que simplificadamente podemos decir que es radial y anular, el Nautilus aborda un cambio de concavidad en la parte dorsal y adopta una curvatura similar tanto en la parte ventral como en la dorsal (Fig. 8) y eso es lo que marca la tendencia a la forma enrollada que adquirirá con posterioridad. La parte dorsal embrionaria es inicialmente casi plana, pero en la denominada constricción de la cicatriz (Fig. 11), coincidiendo con el inicio de la formación de las primeras cámaras y septos, es cuando se produce ese cambio de concavidad que forma el labio interior dorsal (observar en la parte inferior de la Fig. 6-D el piquito que está formándose y en las figuras 12 a 14 cómo se va desarrollando ese labio). Si no se produjera ese cambio de concavidad, irremediablemente la concha se cerraría al intersecar consigo misma. Cuando al crecer, el labio llega a confluir con la parte dorsal inicial de la concha se produce la formación del espacio umbilical u ombligo (Figura 10), momento en el que se completa el primer verticilo y, consecuentemente, la parte ventral inicial de la concha pasa ahora a ser interior a la misma y comienza a actuar como nueva parte dorsal de la cámara habitacional en el segundo verticilo.
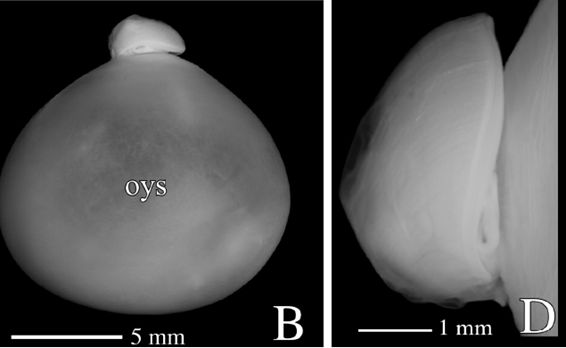
Fig. 6. Embrión de 4 meses del Nautilus pompilius. Imagen B: Perfil con el saco vitelino exterior (oys: outer yolk sac). Imagen D: Detalle de la concha embrionaria. (Procedente de Sasaki, et al., 2010).

Fig. 7 Lapa (Patella). Muséum national d’Histoire naturelle, Paris (France).
 |
 |
|
Fig. 8. Embrión del Nautilus belauesis dentro de la cápsula del huevo (éste ha sido recortado en la parte superior. El embrión contaba con una cámara septal. Procedente de Saunders and Landman (2010: pl. IV, p. xxxiv) |
Fig. 9. Labio dorsal (señalado por la flecha) en la concha del Nautilus en la fase de dos cámaras. (Procedente de Landmam et al.1989). |
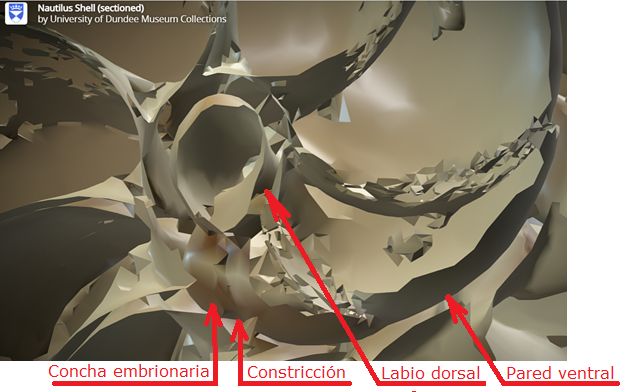
Fig. 10. Detalle del ombligo y de las primeras cámaras. Se observa la concha embrionaria inicial tipo lapa, la constricción que marca el inicio de la pared ventral y el cambio de curvatura en la pared dorsal.
(Procedente de la imagen 3D de la Universidad de Dundee).
Así pues, el perfil de la concha embrionaria preseptal del Nautilus (Fig. 6) puede simplificarse o sintetizarse en dos líneas curvas: la que delimita la parte dorsal y la correspondiente a la ventral (parte izquierda y derecha respectivamente en la figura 6.B). Esta simplificación nos recuerda y hace pensar en la conjunción, antes señalada, entre la espiral longitudinal y la transversal observada en el análisis resumen expuesto en la introducción. Y esa perspectiva se refleja claramente en la sección de dicha concha embrionaria correspondiente a esa fase preseptal (Fig. 11) donde la parte dorsal embrionaria bien podremos catalogarla como el septo cero o inicial (Mutvei & Doguzhaeva, 1997, indican que el protosepto o capa interior sobre la concha inicial es un septo que carece de tubo sifuncular).
Precisamente es en la constricción de la cicatriz (C1 en la Fig. 11) donde la concha adopta la curvatura cordobesa en la pared ventral e inicia el cambio de concavidad en la pared dorsal, iniciándose el labio dorsal. Las figuras 12 a 14 muestran el desarrollo durante la formación de los tres primeros septos.
 |
 |
|
Fig. 11. Desarrollo sucesivo de diferentes capas inorgánicas en la concha preseptal del Nautilus. |
Fig. 12. Sección media dorsoventral de la concha embriónica del Nautilus macromphalus en la etapa del primer septo (S1) y engarce del sifúnculo (Procedente de Tanabe, and Uchiyama, 1997). |
 |
 |
| Fig. 13. Sección media dorsoventral de la concha embriónica del Nautilus belauensis en la fase de dos cámaras (Procedente de Landmam et al.1989. Se corresponde con el ejemplar de la Fig. 9). | Fig. 14. Sección en la concha del Nautilus belauensis en la fase de tres cámaras. Las líneas discontinuas señalan las partes fragmentadas y perdidas al efectuar el corte. (Procedente de Landmam et al.1989). |
En la discusión que realizan en su artículo, Landman et al. (1989) exponen que la concha del Nautilus sufre marcados cambios durante la embriogénesis, en particular indican que durante la etapa de dos y tres cámaras, en el fragmacono, las proporciones que acontecen entre el volumen de las cámaras septales y el de la cámara habitacional son pequeñas y difieren de las alcanzados en etapas ontogénicas posteriores, ello es debido a que la longitud de la cámara habitacional es mucho mayor en la primera etapa vital que en la fase juvenil y adulta. Sin embargo, el perfil de la concha sí se asemeja en todas las fases.
Con la base y los detalles aportados hasta ahora, nos surge el interrogante acerca de: ¿qué acontece con nuestro modelo cordobés uniforme (con igual forma durante todas las etapas vitales) del Nautilus —un modelo bien adaptado a nivel global y macroscópico, como ya hemos mostrado en detalle— si lo confrontamos con esa fase inicial, que biológicamente está indicada como diferenciada? Pues, de partida, lo que ocurre es que se inicia una nueva etapa investigadora, un nuevo camino que recorrer a hombros de estos gigantes biológos, antes explícitamente citados, y de otros muchos más omitidos por economia literaria, pero por supuesto no olvidados, y junto a los de numerosos gigantes matemáticos poder modestamente contribuir a la "Ontogenia matemática del Nautilus". ¡Avancemos, pues!
Para poder adentrarnos en este análisis y contraste, podemos aglutinar todo lo indicado en un recurso interactivo ( ver la escena interactiva 1) que nos permita tener una visión solapada comparativa y que ayude en la conceptualización de lo que acontece, es decir, partir de una agrupación de instantáneas para lograr extraer un continuo descriptivo matemático. Es obvio, de nuevo es necesario indicarlo aunque pueda ser reiterativo, que la superposición de las fotos anteriores mostrará variaciones, algunas debido a que proceden de diferentes ejemplares y especies de Nautilus, otras serán consecuencia de la variabilidad que intrínsecamente acontece en todo espécimen o individuo concreto dentro de las características comunes e identificadoras que comparten y que definen a esa especie; o quizás que estén generadas por posibles desviaciones angulares al realizar los cortes sobre la concha para obtener su sección o perfil. En definitiva, errores comunes e implícitos en cualquier estudio y que aquí, adicionalmente, se ven magnificados gracias a la amplificación visual que nos aporta la técnica. Pero en esa escena, si alguien tiene un empeño cuantificador especial, puede verificar ―con la regla interactiva que se dispone― que nos adentramos en variaciones en el entorno de la décima de milímetro. Estos son detalles con los que se ha de convivir y que han de ponderarse en su justa magnitud para lograr esa visión global y poder encuadrarse en la perspectiva de búsqueda de un modelo o patrón conceptual y así lograr encontrarlo.
A continuación puede interactuar con dicho recurso interactivo y podrá abordar, si lo desea, su investigación personal a través de las diferentes herramientas que ahí se incluyen:
Escena interactiva 1. El modelo cordobes uniforme del Nautilus (revisión)
Nuestra reflexión la dejamos reflejada en la siguiente presentación y puede también usarse como guía a reproducir en la escena interactiva anterior.
Así pues, el análisis realizado nos marca varias necesidades:
- Necesidad de modelar el labio dorsal y el ombligo.
- Necesidad de estudio específico de la concha del Nautilus en el primer verticilo y su ontogenia en general.
- Necesidad de mejorar el modelo de confluencia de septos y paredes en el segundo y tercer verticilo.
En definitiva, contemplamos y concluimos que es obvia la necesidad de abordar la citada "Ontogenia matemática del Nautilus"... Y a ello nos dedicaremos en el siguiente artículo. Para no impacientarse "juegue" con el recurso interactivo aportado y adéntrese en los artículos indicados en la bibliografía descrita.
Bibliografía
MolluscaBase eds. (2021). MolluscaBase. Patella vulgata Linnaeus, 1758. Accessed through: World Register of Marine Species at: http://www.marinespecies.org/aphia.php?p=taxdetails&id=140685 on 2021-08-22
Landman, N. H., Arnold, J. M. and Mutvei, H. 1989: Description of the embryonic shell of Nautilus belauensis. American Museum Novitates, no. 2960, p. 1–16.
Mutvei, H. and Doguzhaeva, L. 1997: Shell ultrastructure and ontogenetic growth in Nautilus pompilius L. (Mollusca: Cephalopoda). Palaeontographica Abteilung A Palaeozoologie–Stratigraphie, vol. 246, p. 33–52.
Sasaki, T., Shigeno S. & Tanabe K. 2010. Anatomy of living Nautilus: Reevaluation of primitiveness and comparison with Coleoidea (2010) in Tanabe, K., Shigeta, Y., Sasaki, T. & Hirano, H. (eds.) 2010. Cephalopods - Present and Past, Tokai University Press, Tokyo, p. 35-66.
Saunders, W. B., and N. H. Landman (eds.) 2010. Nautilus: the biology and paleobiology of a living fossil. (Springer. First edition New Y 1987) Plenum Press, New York.
Tanabe K. & Uchiyama, K. 1997. Development of the Embryonic Shell Structure in Nautilus. The Veliger 40(3): 203-215.
Este obra está bajo una licencia de Creative Commons Reconocimiento-NoComercial-SinObraDerivada 4.0 Internacional

Hace unos años mostramos que la forma y el crecimiento del Nautilus, prototipo de belleza natural y matemática, era neta y completamente cordobés. No obstante, manteniéndose dentro de ese canon, el desarrollo no es uniforme en todas sus etapas vitales y, en particular, en su etapa inicial marca diferencias que son dignas de modelar. Así pues, nos planteamos trasladarles en varios artículos de este blog la ontogenia matemática del Nautilus y le mostraremos que en la primera etapa el modelo se corresponde con un patrón de dos espirales cordobesas con diferente polo y, sin embargo, en la etapa juvenil y adulta éstas comparten el mismo polo. Un mínimo detalle que hay que añadir al buen hacer matemático de este animalito y que da explicación a su acontecer vital, permiténdonos comprender nuevos detalles de su ser.
Quedan invitados a su lectura y quedamos deseosos de sus comentarios y observaciones
Introducción
El Nautilus, usualmente catalogado como un fósil viviente ―si bien Ward (1984) lo encuadra en una estasis aparente[1]―, ha llamado siempre la atención no sólo por la belleza exterior de su concha, quizás no excesivamente destacable frente a la de otros moluscos, sino por aquella que esconde en su interior, ¡que es sorprendente!, y no deja indiferente a nadie. Por ello, ha sido ampliamente estudiada desde el punto de vista paleontológico, zoológico, biológico u ontogénico, pero también en el mundo artístico, el arquitectónico, el ingenieril, el físico-químico y, por supuesto, en el matemático. Incluso ha llegado a ser idealizada en estos entornos científicos, sin olvidar el contexto literario donde también se ha adentrado en lugares preminentes. Aupada como prototipo de belleza ha sido continuamente encajonada de manera forzada en el canon de belleza o proporcionalidad divina o áurea, a pesar de su resistencia y continuo grito de: ¡No!, ¡No soy áureo!, clamor que puede ser inaudible para legos, pero no debería serlo para eruditos. Más fácil hubiera sido haber creado la proporción nautílica, como referente o neotipo de belleza, que con rocambolescas maneras tratar de estirar, contraer o moldear su fina superficie calcárea, pues ello es una suerte imposible de lograr con tan frágil soporte.
En el artículo “Sobre la forma y el crecimiento cordobés del Nautilus pompilius” (Galo, Cabezudo y Fernández, 2016 a) detectamos y expusimos que la espiral logarítmica que se ajusta al perfil de la concha del Nautilus se corresponde con una espiral cordobesa (Fig. 1).

Fig. 1. Ajuste del Nautilus por una espiral logarítmica cordobesa (Galo et al. 2016 b).
Este hallazgo coyuntural que no deja de ser una mera coincidencia numérica, aunque para nosotros muy agradable, con la razón denominada cordobesa y que consecuentemente nos permite asociar su nombre al de Nautilus, hubiera quedado como una curiosidad anecdótica si no fuera porque ahí señalamos y detallamos (ver Fig. 2 y 3) que todo en el Nautilus se muestra cordobés ―sí, nosotros estuvimos silenciosos y atentos y le oímos decir: ¡Qué soy cordobé![2] (ver Fig. 4) ―:
- El rastro calcáreo del sifúnculo (órgano que vacía las cámaras y actúa en la consecución de la flotabilidad del animal) muestra que éste crece siguiendo la forma de esa espiral cordobesa y que es el eje de simetría del teórico cono topológico que forma la cavidad que ha ido construyendo y recorriendo el animal en su ciclo vital.
- Los septos (tabiques que conforman las cavidades de flotación) son arcos de una espiral que es también cordobesa.
- El polo de esos arcos septales se desplaza siguiendo una espiral del mismo tipo, cordobesa, que es la mediatriz de todos los radios vectores de la espiral de la concha.
- Todo punto de la concha o interior es la intersección de dos espirales cordobesas, una longitudinal (tipo a la del perfil de la concha, pero con un factor de escala) y otra transversal (tipo septal).
 |
 |
|
Fig. 2. Espiral que aproxima perfil de la concha (azul), la de los septos (amarilla) y la que describe el polo de los arcos septales (magenta). |
Fig. 3. Detalle de la espirales sobre las que se aproximan los septos: perfil de la concha (azul ),septos (amarilla) y la que describe el polo de los septos (magenta). |
 |
|
Fig. 4. El Nautilus es “cordobé”. |
Así pues, en el artículo citado ” (Galo, Cabezudo y Fernández, 2016 a) se buscó y se logró captar la esencia nautílica matemática a nivel conceptual global: “todo en el Nautilus se muestra cordobés”, y el modelo matemático que ahí se consideró fue único e indiferenciado en todo el crecimiento. Se aplicó la consideración de que dado que el crecimiento del Nautilus es gnomónico, es decir, que mantiene su forma cambiando su escala, se puede hacer una lectura atemporal esperando que el modelo sea aplicable en toda la fase vital desde su germen a su madurez. No obstante, esto, no es más que un útil deseo purista matemático que se mostrará débil para cualquier detenido observador que aborde su confrontación con la realidad, pues somos conscientes por experiencia vital personal que en el desarrollo de un ser vivo acontecen diferentes fases o etapas que introducen matices distintivos dentro de una esencia identificadora que permanece. Y siguiendo esta línea diversificadora, en este artículo lo que se persigue es abordar la ontogenia matemática del Nautilus, desde su periodo embrionario hasta la madurez y determinar cuál es el modelo matemático diferenciado en cada una de las etapas de crecimiento en correlación con los estudios biológicos micro y macroscópicos existentes. Y, de hecho, aquí veremos que se pueden diferenciar tres etapas que esencialmente se corresponden con cada uno de los tres verticilos (un verticilo es una revolución de 360° en el crecimiento en espiral de la concha de un molusco) que se contabilizan en la espiral de la concha de un ejemplar adulto, tres etapas que encierran ciertas diferencias y matices dentro del citado modelo cordobés global. Todo, sin olvidar y teniendo siempre presente, siendo conscientes, que cuando atravesamos el canal que enlaza la discreta yocto-yotta realidad, en la que vivimos, con la idealizada continuidad del mundo matemático siempre surgen interferencias (“aliasing”) que trataremos de mitigar y/o detallar, aunque hay que asumirlas.
La referencia zoológica de la que partiremos es: “Anatomy of living Nautilus: Reevaluation of primitiveness and comparison with Coleoidea” de Sasaki, Shigeno & Tanabe (2010) donde, aportando unas magníficas fotografías, se aborda una revisión de la anatomía del Nautilus, que si bien se centra principalmente en los órganos y partes blandas, incluye también información sobre la forma embrionaria de la concha. Adicionalmente, en la introducción de ese artículo los autores detallan la literatura más significativa al respecto comenzando con el libro de Saunders and Landman (1987) que compila el conocimiento sobre el Nautilus hasta ese año y que posteriormente ha sido actualizado en la edición de 2010. Ladman et al. (1989) analizan la concha en el periodo embrionario y muestran su desarrollo hasta el tercer septo, estudio que se complementa en Mutvei and Doguzhaeva (1997), donde se muestran las curiosas depresiones dorsales septales en el segundo verticilo, y en Tanabe and Uchiyama (1997). Estos artículos nos servirán de soporte, guía y verificación en nuestro análisis matemático. También el modelo virtual tridimensional e interactivo de la concha del Nautilus del “D’Arcy Thompson Zoology Museum”, obtenido mediante escáner, es una magnífica fuente de observación visual interactiva, si bien hay que tener presente y es ostensible que el corte del ejemplar que se ha usado para su digitalización no coincide con el plano medio de simetría. Pueden consultarse otros modelos tridimensionales en Sketchfab.
| Modelo tridimensional de la sección de la concha de un Nautilus pompilius. | |
|
Nautilus Shell (sectioned) by University of Dundee Museum Collections on Sketchfab |
Cortes por planos paralelos al plano medio obtenidos por González-Restrepo (2019) a partir del modelo 3D del Museo Dundee. |
La concha embrionaria y el primer verticilo
Landman et al. (1989) exponen que la forma de la concha del Nautilus sufre marcados cambios durante la embriogénesis y que los perfiles en la etapa de dos y tres cámaras difieren de los alcanzados en etapas ontogénicas posteriores. En nuestro artículo (Galo et al., 2016 a) ya pusimos de manifiesto (Fig. 5) que en el primer verticilo se contabilizaban ocho cámaras, frente a las dieciséis del segundo, es decir, cambia el paso interseptal teórico que en un primer momento es π/4 y posteriormente pasa a ser π/8. Este paso se mantiene también en el tercer verticilo, si bien en esta fase adulta pueden acontecer ciertos cambios morfológicos que influyen en los últimos septos y ése paso puede ser algo menor. En la imagen de la fig. 5 son ocho los septos en el tercer verticilo, pero este número es variable. A pesar de indicar estas diferencias, allí, optamos por presentar un modelo matemático teórico uniforme considerando un crecimiento idéntico en toda la vida del animal y ello permitió fijar los comportamientos globales antes citados como que el sifúnculo sigue el eje de simetría topológico de la cámara habitacional o que los polos de los septos se ubican en la espiral mediana, propiedades que ahora, en un proceso inverso, nos serán de utilidad para analizar y comprender lo que acontece en ese diferenciado primer verticilo. Y ese estudio es el que abordaremos en esta sección donde nos detendremos en la fase embrionaria precameral, en la fase de formación de las primeras cámaras y en lo que acontece en el resto de este primer verticilo.
 |
|
Fig. 5. Recuento de verticilos y cámaras en el Nautilus. |
En el análisis de ese primer verticilo nos adentraremos en un próximo artículo de este blog... no tardará en llegar.
[1] “Rather than being a prime example of a living fossil, the nautiloids may be examples of rapidly speciating organisms that change only slightly during each event, and return to the same form over and over. The result would be apparent stasis, but the actual history would be similar to that of any other rapidly speciating group-except that the net morphologic change over time would be small, rather than large” (Ward, 1984).
[2] En Córdoba (España) la pronunciación de las eses a final de palabra suele ser muy débil y para los oídos no habituados a esa tonalidad, éstas quedan como omitidas.
Bibliografía
Galo J.R., Cabezudo A. y Fernández I.(2016 a) : Sobre la forma y crecimiento cordobés del Nautilus Pompilius. Epsilon, 2016, Vol. 33 (3), nº 94.
Galo J.R., Cabezudo A. y Fernández I.(2016 b) : Recurso interactivo Sobre el crecimiento cordobés del Nautilus Pompilius. Red Educativa Digital Descartes.
González-Restrepo, F. (2019): Cortes del Nautilus a partir de la digitalización 3D del museo Dundee. Red Descartes Colombia.
Landman, N. H., Arnold, J. M. and Mutvei, H. 1989: Description of the embryonic shell of Nautilus belauensis. American Museum Novitates, no. 2960, p. 1–16.
Mutvei, H. and Doguzhaeva, L. 1997: Shell ultrastructure and ontogenetic growth in Nautilus pompilius L. (Mollusca: Cephalopoda). Palaeontographica Abteilung A Palaeozoologie–Stratigraphie, vol. 246, p. 33–52.
Sasaki, T., Shigeno S. & Tanabe K. 2010. Anatomy of living Nautilus: Reevaluation of primitiveness and comparison with Coleoidea (2010) in Tanabe, K., Shigeta, Y., Sasaki, T. & Hirano, H. (eds.) 2010. Cephalopods - Present and Past, Tokai University Press, Tokyo, p. 35-66.
Saunders, W. B., and N. H. Landman (eds.) 2010. Nautilus: the biology and paleobiology of a living fossil. (Springer. First edition New Y 1987) Plenum Press, New York.
Tanabe K. & Uchiyama, K. 1997. Development of the Embryonic Shell Structure in Nautilus. The Veliger 40(3): 203-215.
Ward P. (1984) Is Nautilus a Living Fossil?. In: Eldredge N., Stanley S.M. (eds) Living Fossils. Casebooks in Earth Sciences. Springer, New York, NY. https://doi.org/10.1007/978-1-4613-8271-3_31
Este obra está bajo una licencia de Creative Commons Reconocimiento-NoComercial-SinObraDerivada 4.0 Internacional

Entre los diferentes recursos de la RED para el estudio de ecuaciones cuadráticas, se propone en este artículo una serie de objetos de aprendizaje del subproyecto Prometeo, para el análisis de los diferentes tipos de ecuaciones de 2º grado. Se utilizan diferentes métodos de resolución según se trate de ecuaciones completas o incompletas.
Estas unidades están indicadas para los últimos cursos de la ESO y el bachillerato.
Las unidades seleccionadas pertenecen al grupo Recursos educativos interactivos de matemáticas para el bachillerato, una serie de recursos creados por el Equipo Descartes, que forman parte del proyecto Prometeo, promovido por el Departamento de Educación del Instituto de Matemáticas de la Universidad Nacional Autónoma de México.
Relación de los objetos seleccionados:
Ecuaciones de la forma ax² + c = 0.
Ecuaciones de la forma ax² + bx = 0.
Ecuaciones de la forma a(x+m)² = n.
Ecuaciones de la forma (ax+b)· (cx+d) = 0.
Ecuaciones cuadráticas completas: ax² + bx + c = 0.
En cada unidad se indica, en primer lugar, el procedimiento a seguir para su resolución y análisis del número de soluciones. A continuación se presenta una escena de ejercicios resueltos y finalmente, una escena de ejercicios para practicar con autocorrección.
Para su aplicación en el aula se propone insertar estos recursos en un aula Moodle.
En el siguiente vídeo se puede ver con detalle algunas de las actividades que contienen estos recursos:

Título: Física Mecánica
Sección: iCartesiLibri
Bloque: Física
Unidad: Mecánica
Nivel/Edad: Bachillerato y Universidad (17 años o más)
Idioma: Castellano
Autor: José Ricardo Del Río Quimbayo
Colaboradores: Marco Tulio Mesa Cardona y Jaime Humberto Ramírez Rios
Diseño del libro: Juan Gmo. Rivera Berrío
ISBN: 978-958-52963-6-7
![]() Haz clic aquí para ver una versión en pdf
Haz clic aquí para ver una versión en pdf
![]() Haz clic en la imagen para abrir el recurso
Haz clic en la imagen para abrir el recurso
Puedes encontrar todos los libros interactivos de iCartesiLibri en
https://proyectodescartes.org/iCartesiLibri/index.htm - Ver Créditos
Este material está publicado bajo una licencia:
Creative Commons Reconocimiento-NoComercial-CompartirIgual 4.0 Internacional

Título: Transformo mi función
Sección: Miscelánea
Bloque: Cálculo
Unidad: Operaciones con funciones
Nivel/Edad: Bachillerato y Universidad (17 años o más)
Idioma: Castellano
Autor: José R. Galo Sánchez
![]() Haz clic en la imagen para abrir el recurso
Haz clic en la imagen para abrir el recurso
Puedes encontrar todos los materiales de la Miscelánea en
https://proyectodescartes.org/miscelanea/index.htm - Ver Créditos
Este material está publicado bajo una licencia:

Creative Commons Reconocimiento-NoComercial-SinObraDerivada 4.0 Internacional