Mostrando artículos por etiqueta: bachillerato
.png "Haz click para abrir el recurso")
Título: Curvas y superficies paramétricas (segunda edición)
Sección: iCartesiLibri
Bloque: Geometría
Unidad: Geometría plana y tridimensional
Nivel/Edad: Bachillerato y Universidad (16 años o más)
Idioma: Castellano
Autores: Juan Guillermo Rivera Berrío y Josep Maria Navarro Canut
ISBN: 978-84-18834-35-6
 versión en pdf con enlace a los interactivos
versión en pdf con enlace a los interactivos
![]() Haz clic en la imagen para abrir el recurso
Haz clic en la imagen para abrir el recurso
Puedes encontrar todos los libros interactivos de iCartesiLibri en
https://proyectodescartes.org/iCartesiLibri/index.htm - Ver Créditos
Este material está publicado bajo una licencia:

Creative Commons Reconocimiento-NoComercial-CompartirIgual 4.0 Internacional
.png "Haz click para abrir el recurso")
Título: Integrando con Paco
Sección: iCartesiLibri
Bloque: Análisis matemático
Unidad: Integración
Nivel/Edad: Bachillerato y Universidad (16 años o más)
Idioma: Castellano
Autores: Juan Guillermo Rivera Berrío, y José Román Galo Sánchez
ISBN: 978-84-18834-34-9
versión en pdf con enlace a los interactivos
![]() Haz clic en la imagen para abrir el recurso
Haz clic en la imagen para abrir el recurso
Puedes encontrar todos los libros interactivos de iCartesiLibri en
https://proyectodescartes.org/iCartesiLibri/index.htm - Ver Créditos
Este material está publicado bajo una licencia:
Creative Commons Reconocimiento-NoComercial-CompartirIgual 4.0 Internacional

Esta semana presentamos la unidad Estadística básica, un objeto digital interactivo con actividades de introducción al estudio estadístico, para los cursos de bachillerato y la Universidad.
En esta unidad se introducen las variables estadísticas, las medidas de centralización y de dispersión y se indican los pasos necesarios para la construcción de tablas de frecuencia y diagramas de barras.
La unidad consta de cuatro fases:
Motivación. En esta primera fase se presentan dos vídeos de introducción al tema.
Inicio. Este apartado contiene tres escenas: una primera actividad de clasificación de diferentes tipos de variables, una segunda página con ejercicios para calcular las medidas de centralización y finalmente una escena con ejercicios de cálculo de las medidas de dispersión.
Desarrollo. Incluye actividades de cálculo de medidas estadísticas, elaboración de tablas de frecuencia y diseño de gráficas de barras.
Cierre. Comprende un resumen de los conceptos estudiados y actividades de autoevaluación para consolidar lo aprendido.
La unidad Estadística básica pertenece al proyecto Un_100, un proyecto que agrupa recursos educativos de las áreas de Matemáticas y de Física para los niveles de bachillerato y Universidad. En su elaboración han participado académicos de México, España, Colombia y Chile.

Título: Cálculo diferencial e integral, módulo II.
Sección: iCartesiLibri
Bloque: Análisis matemático
Unidad: Cálculo diferencial
Nivel/Edad: Bachillerato y Universidad (16 años o más)
Idioma: Castellano
Autores: Carlos Alberto Rojas Hincapié
ISBN obra completa: 978-84-18834-30-1
ISBN del volumen: 978-84-18834-33-2
aquí para ver una versión en pdf
![]() Haz clic en la imagen para abrir el recurso
Haz clic en la imagen para abrir el recurso
Puedes encontrar todos los libros interactivos de iCartesiLibri en
https://proyectodescartes.org/iCartesiLibri/index.htm - Ver Créditos
Este material está publicado bajo una licencia:
Creative Commons Reconocimiento-NoComercial-CompartirIgual 4.0 Internacional

Título: Cálculo diferencial e integral, módulo I.
Sección: iCartesiLibri
Bloque: Análisis matemático
Unidad: Cálculo diferencial
Nivel/Edad: Bachillerato y Universidad (16 años o más)
Idioma: Castellano
Autores: Carlos Alberto Rojas Hincapié
ISBN obra completa: 978-84-18834-30-1
ISBN del volumen: 978-84-18834-31-8
![]() Haz clic aquí para ver una versión en pdf
Haz clic aquí para ver una versión en pdf
![]() Haz clic en la imagen para abrir el recurso
Haz clic en la imagen para abrir el recurso
Puedes encontrar todos los libros interactivos de iCartesiLibri en
https://proyectodescartes.org/iCartesiLibri/index.htm - Ver Créditos
Este material está publicado bajo una licencia:
Creative Commons Reconocimiento-NoComercial-CompartirIgual 4.0 Internacional
En nuestro modelo uniforme del Nautilus (Galo et al. 2016) detectamos que los septos son arcos de espirales cordobesas y, adicionalmente, que sus respectivos polos estaban ubicados también en una espiral cordobesa. Ése fue, quizás, el mayor y más novedoso avance logrado en la modelación de esta concha. No obstante, allí, no abordamos la evidente diferencia que acontece entre ellos según las etapas vitales de estos especímenes y, consecuentemente, lo que ocurre también en las cámaras septales que determinan. En especial, esas diferencias se presentan entre el primer verticilo y en los dos restantes. Allí, tampoco profundizamos en el modelado de las intersecciones de los septos con la pared ventral y la dorsal y sólo aventuramos una posible base teórica del fragmacono en base al gnomon de un triángulo cordobés. Ahora, habiendo profundizado en el estudio de la literatura existente sobre la ontogenia biológica del Nautilus y también en el análisis matemático del modelo propuesto entonces, es el momento de adentrarnos en un modelo matemático diferenciado por fases, es decir, de abordar la ontogenia matemática de los septos.
Siguiendo lo indicado en los artículos anteriores de esta serie (puede consultar: I, II, III y IV), y en particular en lo relativo a la modelación de la pared dorsal y ventral de la sección sagital de la concha, partimos de una base o fundamento primordial que es el que justifica y explica el distinto comportamiento de los septos entre el primer verticilo y los siguientes. En el primero, ambas paredes se corresponden con arcos de dos espirales cordobesas que tienen diferente polo, sin embargo, en el segundo y en el tercer verticilo las espirales de ambas paredes son copolares, tienen el mismo polo, siendo realmente arcos de una única espiral con un retardo angular entre ambas de 2π. Así pues, dado que la complejidad es mayor en el primer verticilo vamos a proceder en orden cronológico inverso y analizaremos en primer lugar el segundo y tercer verticilo y posteriormente el primero.
Los septos en el segundo y tercer verticilo
La pared ventral en el segundo y tercer verticilo viene dada por
 (20)
(20)
donde θi es el ángulo que marca el inicio de la concha embrionaria y θf la terminación de la misma delimitando la boca de la concha.
Y en el mismo instante[1] angular vital θ, la pared dorsal sería:
 (21)
(21)
La espiral que contiene a los polos de los septos es:
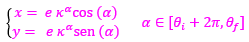 (22)
(22)
donde e es un factor de escala o de retardo en la espiral cordobesa [2].
Y cada septo, ver (11 en IV), es un arco de una espiral:
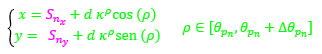 (23)
(23)
donde 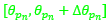 determina la amplitud angular del arco de esa espiral que comprende el septo n-ésimo; ρ determina unívocamente cada uno de los puntos de dicho septo, pero todos ellos se corresponden con un mismo instante vital[3]; d es un factor de escala o de retardo a determinar en el modelo; y
determina la amplitud angular del arco de esa espiral que comprende el septo n-ésimo; ρ determina unívocamente cada uno de los puntos de dicho septo, pero todos ellos se corresponden con un mismo instante vital[3]; d es un factor de escala o de retardo a determinar en el modelo; y 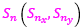 es el polo de la espiral que incluye a ese arco septal n-ésimo y que perteneciendo a la espiral (22) quedará determinado por un valor αn.
es el polo de la espiral que incluye a ese arco septal n-ésimo y que perteneciendo a la espiral (22) quedará determinado por un valor αn.
La aplicación en el modelo de la que hemos denominado invariante tercera (tangencialidad entre la pared ventral y las paredes de los septos) nos puede llevar a determinar los parámetros antes citados. De partida:
- Toda espiral logarítmica es equiangular, así pues, en cualquier punto de la espiral la recta tangente y el radio vector forman siempre un mismo ángulo ψ. Éste es característico de cada tipo espiral y depende sólo de la base b que la define, siendo
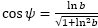 . En el caso de una espiral cordobesa este ángulo es ψ ≃ 80,32º, al ser la base logarítmica o exponencial que la define κ = 1,185580...
. En el caso de una espiral cordobesa este ángulo es ψ ≃ 80,32º, al ser la base logarítmica o exponencial que la define κ = 1,185580... - Al ser tangentes la espiral ventral y la septal, ambas comparten la misma recta tangente. Y dado que ambas espirales son cordobesas entonces, consecuentemente, los radios vectores de ambas han de estar también en la misma recta, porque ambos han de formar el mismo ángulo con la tangente común.
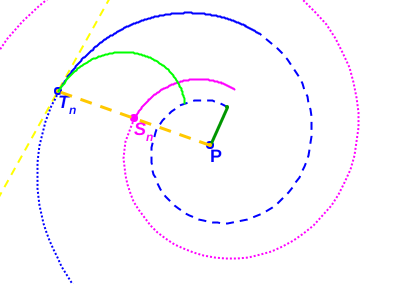
(i) Espiral azul discontinua: pared dorsal en el segundo verticilo. (ii) Espiral azul continua y de puntos: pared ventral en el segundo verticilo.
(iii) Espiral magenta: espiral de los polos de los septos. (iv) Tn: punto de tangencia septo y pared ventral; Sn: polo del septo y P: polo común de la espiral dorsal, de la ventral y de la de los polos de los septos.
Fig. 40. Tangencialidad de los septos con la pared ventral.
Por tanto (ver Fig. 40), si Tn es el punto de tangencia del n-ésimo septo (con n>8, pues en el primer verticilo hay ocho septos), Sn es el polo de éste y P el polo de la pared ventral, tenemos que:
- Al ser Tn un punto de la espiral ventral, entonces:
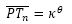 (24)
(24)
para algún θ.
- Al ser Sn un punto de la espiral de los polos de los septos, por (22),
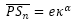 (25)
(25)
donde α = θ al estar alineados P, Sn, y Tn y ser P el polo común a la espiral ventral (20) y a la espiral de los polos de los septos (22).
- Al ser Tn un punto del septo n-ésimo
![]() (26)
(26)
para algún valor de d y ρ.
Y dado que
![]() (27)
(27)
de las relaciones anteriores, (24) a (26), obtenemos que:
![]() . (28)
. (28)
Expresando en (28) d = d' κθ - ρ, es decir, considerando que ρ es un ángulo de retardo, tenemos:
![]() (29)
(29)
Y de ahí
d' = 1 - e. (30)
En Galo et al. (2016) detectamos que en la espiral de los polos de los septos (22) e ≃ 0,5 e igual acontecía para la espiral que da forma a los arcos de los septos. Aquí la relación obtenida en (30) conduce a considerar que e = 0,5 (exactamente ese valor[4]), pues en ese caso también es d' = 0.5, y consecuentemente la espiral correspondiente a un determinado arco septal se obtiene sin más que realizar una traslación de la espiral de los polos para que el polo de ésta coincida con el polo de dicho septo. (ver fig. 41).
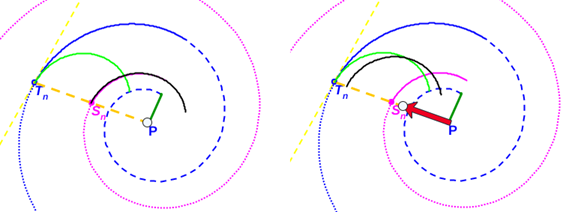
Fig. 41. Obtención de un arco septal como traslación de un arco de la espiral de los polos.
Intersección de la pared dorsal y los septos en el segundo y tercer verticilo
Centrémonos ahora en la determinación de la intersección de los arcos de los septos con la pared dorsal y la amplitud de estos.
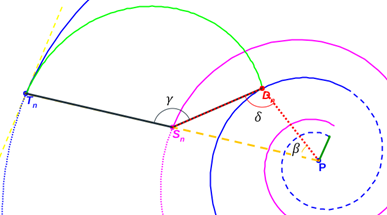
Fig. 42. Parámetros que definen los septos en el segundo y tercer verticilo.
Para el septo n-ésimo, según la denominación de los ángulos reflejados en la fig. 42 y fijado el valor de e = 0,5, por (25) tenemos que:
![]() (31)
(31)
El punto Dn, intersección de ese septo con la pared dorsal, por pertenecer a ella y según (21) verifica que
![]() (32)
(32)
y, a su vez, por pertenecer al arco del septo:
![]() (33)
(33)
- Aplicando el teorema del coseno en el triángulo de vértices P, Sn y Dn:
![]() (34)
(34)
y considerando las expresiones (31), (32) y (33) llegamos a la igualdad:
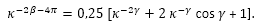 (35)
(35)
- Aplicando el teorema del coseno en el triángulo de vértices Tn, Sn y Dn, y teniendo en consideración que
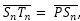 obtenemos que:
obtenemos que:
![]() (36)
(36)
Y puesto que las coordenadas de los puntos que intervienen en esa igualdad son:
![]()
se tiene que:
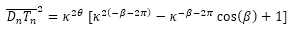 (37)
(37)
Y teniendo en consideración (31) y (33)
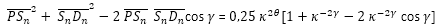 (38)
(38)
Por tanto, la igualdad (36) queda expresada como:
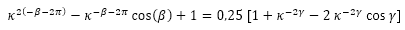 (39)
(39)
A partir de (35) y (39) tenemos un sistema de dos ecuaciones que nos relaciona al ángulo γ (amplitud del arco del septo), con el β (retardo del punto de intersección dorsal del septo Dn, respecto al punto de intersección ventral Tn). Este sistema puede reescribirse como:
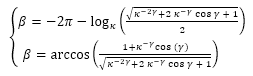 (40)
(40)
Es decir,
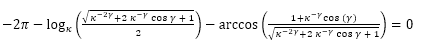 (41)
(41)
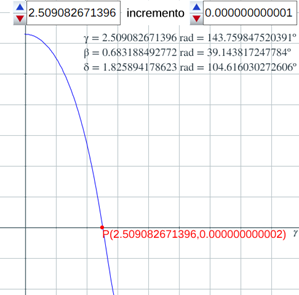
Escena interactiva 5. Determinación numérica de la amplitud del septo.
Pulse sobre la imagen para interactuar libremente con ella.
La resolución numérica de la ecuación (41) (puede observarse en la escena interactiva 5, donde la gráfica en azul se corresponde con la función en la variable γ, definida por la expresión del miembro de la izquierda en (41) con 0 ≤ γ ≤π) nos permite determinar:
- La amplitud del septo
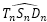 : γ = 2,5090... radianes ≃ 143,76º.
: γ = 2,5090... radianes ≃ 143,76º. - El desplazamiento entre la intersección dorsal y la ventral
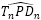 : β = 0,6831... radianes ≃ 39,14º.
: β = 0,6831... radianes ≃ 39,14º. - El ángulo entre los radios vectores
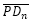 y
y 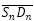 :
: 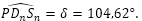
Apoyándonos en que en una espiral cordobesa el ángulo que forma el radio vector con la recta tangente es de 80,32 º tenemos que (ver Fig. 43) el ángulo que forma la recta tangente a la pared dorsal en Dn con la recta tangente al septo en ese mismo punto es de 75,38º, es decir el septo no interseca a la pared dorsal perpendicularmente, si no formando con respecto a esa perpendicular un ángulo de 14,62º. Eso concuerda con lo indicado por Mutvei & Doguzhaeva (1997), que ya reflejamos en la figura 20, y la depresión septal dorsal en el área media ―sección o corte que es el que estamos analizando en este estudio― lo que hace es corregir dicha desviación respecto a la perpendicular buscando aportar y lograr, quizás, una mayor consistencia (eso es lo que puede interpretarse de este hecho aportado por la matemática).
Este comportamiento teórico es el mismo tanto en el segundo como en el tercer verticilo, pues en ambos casos la pared dorsal y ventral comparten el mismo polo. En el tercer verticilo, esa depresión dorsal parece ser menos notable, al menos aparentemente, si bien sí pueden visualizarse o intuirse (interactuar por ejemplo con la digitalización del Nautilus del Museo Dundee ―figura 22―). Matemáticamente no hay diferencia.
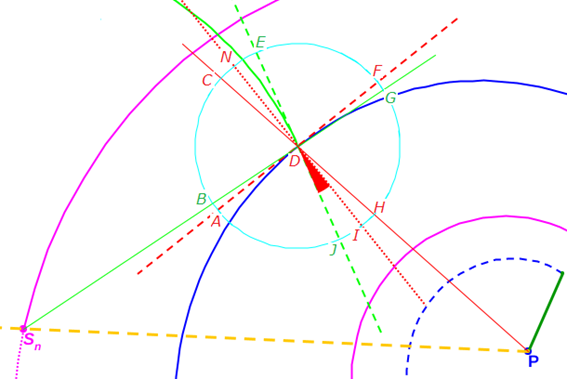

Fig. 43. Ángulo de incidencia entre septo y pared dorsal en el segundo y tercer verticilo.
Ecuaciones del modelo
Así pues, en estos dos verticilos los arcos de los septos tienen por ecuación (23), donde para cada n ≥ 8 (en el primer verticilo hay ocho septos y el octavo da inicio al segundo) tenemos que:
- d = 0,5.
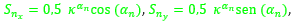 siendo
siendo 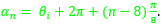
- ρ ∈ [ αn - γ, αn ], con γ = 2,5090...
Los puntos de tangencia son: Tn ( ![]() ) y los de intersección con la pared dorsal pueden escribirse:
) y los de intersección con la pared dorsal pueden escribirse:
- como punto de la pared dorsal: Dn (
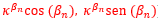 ), donde denotamos βn = αn - 2π - β con β = 0,6831.
), donde denotamos βn = αn - 2π - β con β = 0,6831. - como punto del arco de septo:
 ,
, 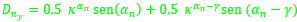 .
.
Síntesis
La introducción de la tangencialidad entre la pared ventral y los septos nos ha permitido lograr la modelación matemática de la sección sagital del Nautilus en el segundo y tercer verticilo y acentuar el caracter cordobés de sus elementos. En la escena interactiva 6, podemos observar e interactuar con este modelo.
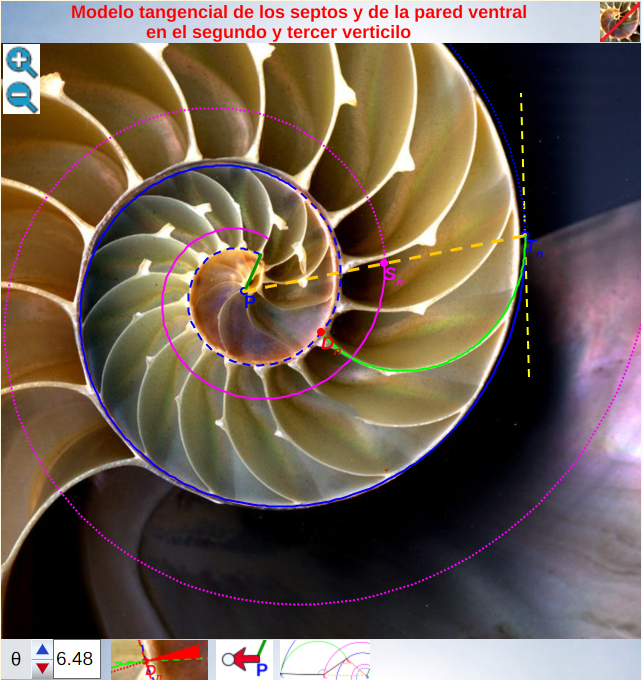
Escena interactiva 6. Modelo tangencial de los septos y de la pared ventral en el segundo y tercer verticilo.
Pulse sobre la imagen para interactuar libremente con ella.
El camino seguido en el análisis anterior nos puede servir de guía para abordar el estudio de los septos en ese primer verticilo tan especial. Especial por ser la pared dorsal y la ventral espirales cordobesas con distinto polo y, como veremos, por ser ésta la causa esencial de esas cámaras diferentes y esos septos variables. Un estudio que considero es interesante y matemáticamente bonito. Pero siento dejarles con la miel en los labios ya que lo dejaré para un nuevo artículo... espero no tenerles en vilo mucho tiempo.
Bibliografía
Galo J.R., Cabezudo A. y Fernández I.(2016) : Sobre la forma y crecimiento cordobés del Nautilus Pompilius. Epsilon, 2016, Vol. 33 (3), nº 94.
Mutvei, H. and Doguzhaeva, L. (1997): Shell ultrastructure and ontogenetic growth in Nautilus pompilius L. (Mollusca: Cephalopoda). Palaeontographica Abteilung A Palaeozoologie–Stratigraphie, vol. 246, p. 33–52.
Ward, P. (1979). Cameral liquid in Nautilus and ammonites. Paleobiology, 5(1), pp. 40-49.
Ward, P., Greenwald, L., & Magnier, Y. (1981). The chamber formation cycle in Nautilus macromphalus. Paleobiology, 7(4), 481-493. doi:10.1017/S0094837300025537
[1] Al no tener una referencia temporal del crecimiento del Nautilus, sólo podemos señalar un mismo instante vital teórico mediante el uso de una amplitud angular común. De esta manera establecemos momentos, atemporales, en la que se han de dar coincidencias vitales. En este caso, para un valor fijado de θ, conocemos el punto de la pared ventral y el punto de la pared dorsal que están relacionados entre sí.
[2] En estos verticilos, en el modelo uniforme se detectó que e ≃ 0,5, que es un valor próximo al valor medio del factor correspondiente al sifúnculo y al del de la pared dorsal ―Galo et al., 2016―.
[3] Para conocer el proceso de formación de las cámaras de los nautilos podemos acudir a lo estudiado y analizado por Ward, Greenwald y Magnier (1981) en su artículo “The chamber formation cycle in Nautilus macromphalus”. Estos autores basan su estudio en la observación radiográfica (ver figura 26) de diferentes ejemplares en distintos momentos y, así, pueden analizar las variaciones que acontecen y realizar mediciones que llevan a plantear un crecimiento periódico que comprende tres fases:
- Formación de una cresta mural en la posición que ocupará el nuevo septo. Esta cresta es una delgada banda anular interna de carbonato cálcico.
- Desplazamiento hacia delante del manto septal para ubicarse a la altura de la cresta mural y ajustarse a ella. Inicio del proceso de calcificación del nuevo septo. También el sifúnculo comienza a calcificar un anillo de conexión en el interior de la nueva cámara uniendo el septo anterior y el nuevo. Durante esta fase la nueva cámara está llena de líquido cameral (Ward, 1979) y no acontece ningún vaciado de la misma, pero ese vaciado sí continúa en las cámaras anteriores.
- Vaciado del líquido de la nueva cámara, que se inicia cuando el nuevo tabique ha alcanzado de un tercio a dos tercios de su espesor final. Este vaciado se denomina acoplado pues el líquido está en contacto con el anillo de conexión sifuncular. En esta fase el tabique septal sigue construyéndose, engrosándose, finalizando este proceso cuando el volumen del líquido vaciado es aproximadamente el 50% y ya no está en contacto con el anillo sifuncular, momento en el que se pasa a un proceso de vaciado desacoplado y comienza la formación de una nueva cresta mural y, consecuentemente, un nuevo ciclo.
Durante el ciclo de formación de una cámara, el crecimiento de la concha exterior parece ser que es continuo, pero hay una correlación inversa entre el porcentaje de líquido que se ha vaciado en la última cámara construida y la amplitud angular de la cámara habitacional. A medida que la cámara septal está más vacía la cámara habitacional es mayor y viceversa. Esta relación logra mantener la flotabilidad ya que cuando la nueva cámara está más llena de líquido el peso de la concha en la zona habitacional es menor y a medida que decrementa el líquido aumenta la amplitud de la zona habitacional. El inicio de cada cámara representa un punto crítico para la flotabilidad global, pero esto se compensa con el vaciado desacoplado que sigue aconteciendo en las cámaras anteriores.
[4] También se apuntó la posibilidad de que ese valor correspondiera a la espiral intermedia entre la del sifúnculo y la pared dorsal, es decir, ![]() , pero en este caso d' ≠ e, es decir las dos espirales citadas son diferentes. De ahí que optemos por el valor e = 0.5.
, pero en este caso d' ≠ e, es decir las dos espirales citadas son diferentes. De ahí que optemos por el valor e = 0.5.

Este obra está bajo una licencia de Creative Commons Reconocimiento-NoComercial-SinObraDerivada 4.0 Internacional

Título: Introducción a la electrónica. Componentes y aplicaciones.
Sección: iCartesiLibri
Bloque: Ingeniería
Unidad: Electrónica
Nivel/Edad: Bachillerato y universidad (16 años o más)
Idioma: Castellano
Autores: Oscar Ignacio Botero Henao, Jesús Ignacio Calle Pérez, Diego Hernando Orozco Gómez y Sergio Hernando Ruiz
ISBN: 978-84-18834-24-0
![]() Haz clic aquí para ver una versión en pdf
Haz clic aquí para ver una versión en pdf
![]() Haz clic en la imagen para abrir el recurso
Haz clic en la imagen para abrir el recurso
Puedes encontrar todos los libros interactivos de iCartesiLibri en
https://proyectodescartes.org/iCartesiLibri/index.htm - Ver Créditos
Este material está publicado bajo una licencia:
Creative Commons Reconocimiento-NoComercial-CompartirIgual 4.0 Internacional

Entre los múltiples proyectos de la RED, encontramos una serie de recursos educativos que se pueden modificar de forma sencilla, permitiendo al profesorado crear nuevos materiales sin necesidad de ser desarrollador de escenas Descartes. Nos referimos a los materiales del subproyecto Plantillas con Descartes-JS.
En este artículo vamos a mostrar los diferentes modelos de actividades que se pueden realizar con estos recursos y veremos algunos materiales elaborados con dichas plantillas.
El contenido de este subproyecto se estructura siguiendo las opciones del menú lateral de la web del subproyecto:
Descripcion. Explicación del subproyecto.
Materiales. Índice de plantillas agrupadas en objetos interactivos y libros interactivos.
-
-
- Objetos interactivos. Diferentes plantillas para generar juegos, actividades de selección, asociación, emparejamientos, puzles… Para cada plantilla se dispone del enlace a un modelo del recurso que sirve de base para realizar nuevo material, el documento en pdf para acceder a la guía con indicaciones para modificar la plantilla y un archivo comprimido para descargar la plantilla.
- Libros interactivos. Una selección de diferentes plantillas para crear libros interactivos.
- Contenedores. Cada contenedor engloba diferentes tipos de recursos y plantillas en una misma presentación.
-
Tutoriales. Documentos y vídeos para guiar al profesorado en la elaboración de sus propios recursos.
Aplicaciones. Para su aplicación directa en el aula, en este apartado se dispone de diferentes unidades realizadas por otros usuarios a partir de las plantillas. Estos objetos interactivos están agrupados por áreas y dentro de cada área se encuentran clasificados por temas o por niveles.
En el siguiente vídeo se presenta la web del subproyecto plantillas y se muestra, a modo de ejemplo, una serie de actividades de lengua para primero de primaria.

Título: Estructuras en madera. Nociones básicas.
Sección: iCartesiLibri
Bloque: Ingeniería
Unidad: Estructuras y materiales
Nivel/Edad: Bachillerato y universidad (16 años o más)
Idioma: Castellano
Autores: John Jairo García Mora y Sonia Jaquelliny Moreno Jiménez
ISBN: 978-84-18834-22-6
![]() Haz clic en la imagen para abrir el recurso
Haz clic en la imagen para abrir el recurso
Puedes encontrar todos los libros interactivos de iCartesiLibri en
https://proyectodescartes.org/iCartesiLibri/index.htm - Ver Créditos
Este material está publicado bajo una licencia:
Creative Commons Reconocimiento-NoComercial-CompartirIgual 4.0 Internacional

Título: Ejercicios de Trigonometría. Procesos, métodos y actitudes.
Sección: iCartesiLibri
Bloque: Geometría
Unidad: Trigonometría
Nivel/Edad: Bachillerato y universidad (16 años o más)
Idioma: Castellano
Autor: José Antonio Salgueiro González
ISBN: 978-84-18834-21-9
![]() Haz clic en la imagen para abrir el recurso
Haz clic en la imagen para abrir el recurso
Puedes encontrar todos los libros interactivos de iCartesiLibri en
https://proyectodescartes.org/iCartesiLibri/index.htm - Ver Créditos
Este material está publicado bajo una licencia:
Creative Commons Reconocimiento-NoComercial-CompartirIgual 4.0 Internacional






