Comunicación DescartesJS-GeoGebra Segunda Edición

Título: Comunicación DescartesJS-GeoGebra. Segunda edición
Sección: iCartesiLibri
Bloque: Formación
Unidad: Formación en Descartes JS
Nivel/Edad: Bachillerato y Universidad (16 años o más)
Idioma: Castellano
Autores: Juan Guillermo Rivera Berrío y Joel Espinosa Longi
ISBN: 978-84-18834-45-5
![]() Haz clic aquí para ver una versión en pdf
Haz clic aquí para ver una versión en pdf
![]() Haz clic en la imagen para abrir el recurso
Haz clic en la imagen para abrir el recurso
Puedes encontrar todos los libros interactivos de iCartesiLibri en
https://proyectodescartes.org/iCartesiLibri/index.htm - Ver Créditos
Este material está publicado bajo una licencia:

Creative Commons Reconocimiento-NoComercial-CompartirIgual 4.0 Internacional
Estadística, probabilidad e inferencia
.png "Haz click para abrir el recurso")
Título: Estadística, probabilidad e inferencia
Sección: iCartesiLibri
Bloque: Estadística y probabilidad
Unidad: Estadística, probabilidad e inferencia
Nivel/Edad: Bachillerato y Universidad (16 años o más)
Idioma: Castellano
Autores: Juan Jesús Cañas Escamilla y José R. Galo Sánchez
ISBN: 978-84-18834-44-8
![]() Haz clic aquí para ver una versión en pdf
Haz clic aquí para ver una versión en pdf
![]() Haz clic en la imagen para abrir el recurso
Haz clic en la imagen para abrir el recurso
Puedes encontrar todos los libros interactivos de iCartesiLibri en
https://proyectodescartes.org/iCartesiLibri/index.htm - Ver Créditos
Este material está publicado bajo una licencia:
Creative Commons Reconocimiento-NoComercial-CompartirIgual 4.0 Internacional
Octava sesión de la 3ª edición del curso "Edición de libros interactivos"
Octava sesión (1 de julio de 2022)
|
1A.
Material previo a esta sesión:
|
Sitios singulares del Nautilus en el primer verticilo

Título: Sitios singulares del Nautilus en el primer verticilo
Sección: Miscelánea
Bloque: Geometría
Unidad: Geometría plana
Nivel/Edad: Universidad (18 años o más)
Idioma: Castellano
Autoría: José R. Galo Sánchez
![]() Haz clic en la imagen para abrir el recurso
Haz clic en la imagen para abrir el recurso
Puedes encontrar todos los materiales de la Miscelánea en
https://proyectodescartes.org/miscelanea/index.htm - Ver Créditos
Este material está publicado bajo una licencia:
Creative Commons Reconocimiento-NoComercial-SinObraDerivada 4.0 Internacional
Ontogenia matemática del Nautilus (VII)

En el primer verticilo del Nautilus encontramos algunos sitios que podemos catalogar como singulares, pues se muestran como específicos respecto a su entorno o que aparentemente se salen de la uniformidad cordobesa que hemos ido detectando y desgranando en los artículos publicados anteriormente sobre la "Ontogenia matemática del Nautilus". Pero, hasta en estas situaciones, la proporción cordobesa y la espiral cordobesa aparece sin más que rascar matemáticamente en la concha del Nautilus, por supuesto con gran mimo.
Esos sitios singulares son:
- La concha embrionaria.
- El sifúnculo en la segunda cámara septal y su relación con la ampliación del fragmacono en el primer septo.
- La confluencia de la pared dorsal con la concha embrionaria en la fase de transición entre el primer y segundo verticilo (cámaras octava, novena y décima).
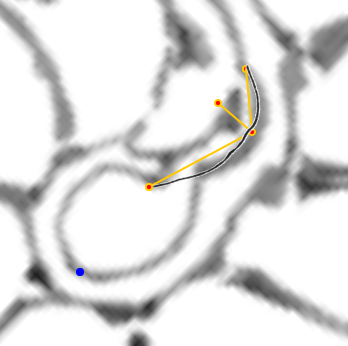 |
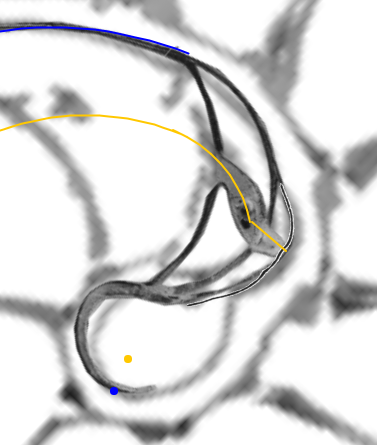 |
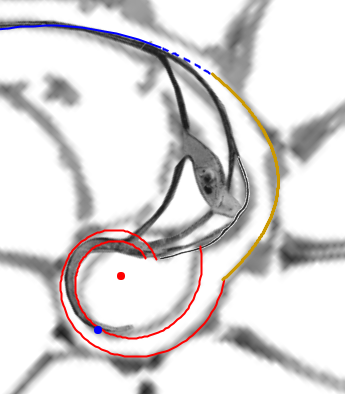 |
| La concha embrionaria | El sifúnculo en la segunda cámara septal y la ampliación del fragmacono |
La pared dorsal en la transición entre el primer y segundo verticilo |
En este artículo analizamos estas singularidades y encuadramos su modelado en el contexto cordobés del Nautilus. Para ello, acudimos y aplicamos la invariante enunciada desde el inicio y que reiteradamente hemos ido aplicando en toda la modelación: "Todo punto interior a la concha o sobre ella se obtiene como la intersección de dos espirales cordobesas, una longitudinal similar a la ventral y otra transversal similar a la septal". La siguiente escena interactiva ha servido de base para el análisis de estos sitios singulares.
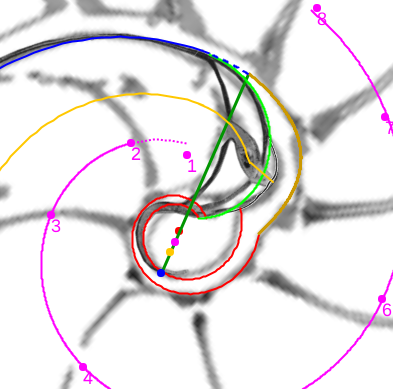
Modelando los sitios singulares del Nautilus en el primer verticilo
Pulsad sobre la imagen para acceder a la escena
En el siguiente pdf (o desde este enlace) tienen desarrollados los contenidos de este artículo
Ontogenia matemática del Nautilus VII
Aquí, hemos dado continuidad a los artículos anteriores (I, II, III, IV, V y VI) y por fin, creo, he cumplido mis deberes, si bien no seré yo quien excluya la posibilidad de que puedan surgir nuevas cuestiones, pues eso es lo que ha ido aconteciendo a lo largo de estos meses en los que les he ido relatando mi investigación a través de este blog de RED Descartes. Pero, independientemente de que afloren nuevas cuestiones o no, sí les indico que éste no será este el último artículo de esta serie, tengo que tratar de completarla adecuadamente y, por tanto, estimo necesario recopilar todo lo expuesto en una escena interactiva de Descartes o en una animación o en un gif animado o en un recurso similar o... y así mostrar la ontogenia matemática del Nautilus en un hilo temporal que recoja su crecimiento, si bien real y paradójicamente éste ha de ser atemporal porque en la literatura existente los datos relativos a los tiempos en el crecimiento de la concha son muy genéricos o yo no he logrado localizarlos. En la siguiente animación reflejamos el modelo uniforme del Nautilus y próximamente podremos aportar el modelo ontogénico (de nuevo les pido un poquito más de paciencia, no nos demos un atracón que pueda conducir a una indigestión).

Modelo uniforme del Nautilus
Y también tendremos que desarrollar alguna cosita en 3D ¿no les parece?... Hasta pronto...

Este obra está bajo una licencia de Creative Commons Reconocimiento-NoComercial-SinObraDerivada 4.0 Internacional






