
La esencia del Nautilus es ostensiblemente cordobesa, pero al analizar su ontogenia biológica se detecta un comportamiento diferente en cada uno de los tres verticilos que pueden contabilizarse en la concha de todo animal adulto. Esta diferencia es especialmente manifiesta y significativa en el primero de ellos. En este artículo, nos ponemos nuestras gafas matemáticas para tratar de localizar la causa y así poder mejorar el modelo matemático uniforme que previamente habíamos compartido. Aquí, nos adentramos plenamente en la ontogenia matemática del Nautilus. Toda espera queda satisfecha para quienes son pacientes y están interesados.
En los artículos previos (Ontogenia I y Ontogenia II) llegamos a detectar la necesidad de una revisión del modelo cordobés uniforme del Nautilus, en especial en el primer verticilo. Y para lograr modelar la ontogenia matemática citada, tendremos que apoyarnos en la ontogenia biológica detectando en ésta los diferentes detalles y particularidades que determinan las características esenciales en cada momento y etapa, y tendremos que concretar cada una de ellas para su adecuada traslación al modelo matemático final. Esto es lo que abordaremos a continuación.
Observando la ontogenia biológica del Nautilus con gafas matemáticas
Concha embrionaria
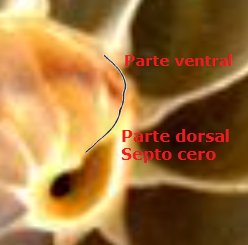
La concha embrionaria inicial, aquella que no tiene ninguna cámara y septo, y de ahí que se le denomine preseptal, tiene aspecto de lapa (Figuras 7 y 10). Su perfil (imagen A de la Fig. 11 y Fig. 15) queda modelado por dos curvas: una que se corresponde con la parte ventral, aquella que tendrá continuidad en la pared ventral de la concha, y otra dorsal que podría considerarse como el septo inicial o septo cero (Mutvei & Doguzhaeva, 1997). Esta característica primigenia se traslada y mantiene permanente en todas la etapas vitales, es un invariante, pues como conocemos se verifica que todo punto interior a la concha o sobre ella se obtiene como la intersección de dos curvas espirales, una longitudinal similar a la ventral y otra transversal similar a la septal.
Curvatura ventral
La constricción de la cicatriz, coincidente con la intersección de la concha embrionaria con el primer septo, es la que marca el inicio del patrón de crecimiento correspondiente a la sección de la concha ventral. A partir de la imagen correspondiente a la fase del primer septo (ver Fig. 16) podemos observar claramente la curvatura de la parte ventral y verificar cómo ésta se ajusta a una espiral cordobesa, ajuste que permanece durante toda la vida del animal (Fig. 17). Consecuentemente, ésta es una característica intrínseca y permanente, un segundo invariante, en toda la ontogenia del Nautilus desde el periodo embrionario septal a la madurez.
 |
 |
 |
| Fig. 15. Perfil de la concha embrionaria o preseptal | Fig. 16. Ajuste de la pared ventral en la fase del primer septo. C1: Constricción |
Fig. 17. El perfil ventral es cordobés en toda la etapa vitall |
Tangencialidad de los septos con la pared ventral de la concha
En las imágenes de las figuras 12, 13 y 14 puede intuirse que las paredes septales confluyen con la pared ventral de la concha tangencialmente, lo cual a priori parece lógico que acontezca para que el cuerpo del animal pueda asentarse de manera uniforme en la zona de unión de ambas paredes. Eso se visualiza claramente a nivel microscópico (Fig. 18), donde en la parte apical (parte izquierda en esa imagen) también puede observarse un cemento de unión entre ambas paredes que a nivel macroscópico difumina esta tangencialidad y puede inducir a interpretar inadecuadamente la forma en que ambas capas confluyen. Esta tangencialidad se da en toda la ontogenia del Nautilus, tercer invariante, y necesitamos destacarla y fijarla porque es básica para el modelo matemático buscado.
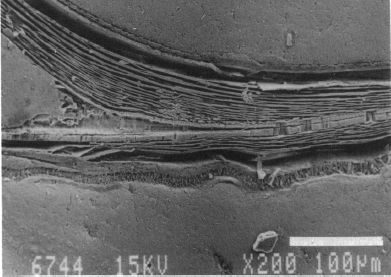
Fig. 18. Detalle de la unión del segundo septo y la parte ventral de la concha en la fase de dos cámaras. Ampliación de la imagen de la Fig. 13. Entre ambas paredes en la parte apical se observa un cemento de unión. (Procedente de Landmam et al.1989).
Incidencia de los septos con la pared dorsal de la concha
La modelación de la incidencia de los septos con la pared dorsal requiere un análisis detallado ya que presenta variaciones destacables en cada verticilo y, por tanto, habrá que localizar la causa de las mismas. Así pues, observando la unión de los dos primeros septos con dicha pared dorsal (Fig. 19) podemos constatar que aquí no acontece, no se reproduce, la indicada tangencialidad ventral y se detecta cemento de unión en la parte apical (en esta imagen ubicada a la derecha) y en la adoral (izquierda). Matemáticamente, si partimos de que la forma del septo sigue una determinada curva prefijada (en el modelo global uniforme indicamos, mostramos y asumimos que es un arco de espiral) y considerando la tangencialidad indicada de éste con la pared ventral, entonces se concluye que la incidencia que acontece con la pared dorsal viene dada, está preestablecida, es forzada y tiene que asentarse según el ángulo que corresponda en cada momento, y quizás ello es lo que constructivamente obliga a que la unión se tenga que reforzar para obtener una consistencia adecuada (de ahí el cemento indicado). Algebraicamente, detectada la forma de la de la pared dorsal, la determinanción del ángulo y punto de incidencia quedan reducidos a intersecar la ecuación del septo con la de la pared dorsal, y eso es lo que haremos en su momento y en cada una de las fases del crecimiento, las cuales hemos asociado a cada verticilo.

Fig. 19. Detalle de la unión de los dos primeros septos (1 y 2) y la parte dorsal de la concha en la fase de dos cámaras. Ampliación de la imagen de la Fig. 13. Entre ambas paredes se observa un cemento de unión. (Procedente de Landmam et al.1989).
Adicionalmente, dado que la parte dorsal de la concha es diferente en el primer verticilo (lo analizaremos posteriormente en un apartado específico), la amplitud del arco del septo parece ser variable y ,si es así, tendrá que acortarse o alargarse hasta encontrar el punto de apoyo necesario para el cierre de cada cámara. Acorde con ello, el ángulo de incidencia observado parece ser diferente en los septos del primer verticilo (observar detenidamente la Fig. 20, imagen A). En esa misma figura se observa que en el segundo verticilo, en el área dorsal media (mda), son ostensibles las depresiones septales dorsales (dsd), ver detalle de su forma en las imágenes B, C y D, y éstas se vuelven menos notables en el tercer verticilo. La ontogenia biológica parece mostrarse esquiva con la matemática o simplemente gusta de marcarnos retos adicionales sobre los que dilucidar.
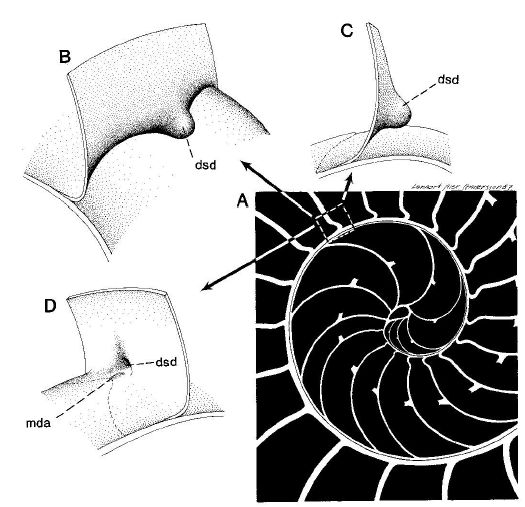
Fig. 20. Sección de la concha del Nautilus. Diferentes vistas de la depresión septal dorsal (dsd) en el área dorsal media (mda). (Procedente de Mutvei & Doguzhaeva, L. 1997).
Labio dorsal y ombligo
Como indicamos con antelación (ver la figura 10) es en la constricción dorsal, que es también el punto de incidencia del primer septo, donde el perfil de la concha en su parte dorsal cambia su concavidad y adopta la misma que la pared ventral, si no fuera así, la cavidad se cerraría. Y al confluir la pared dorsal de este primer verticilo con la concha embrionaria preseptal es como se forma el ombligo.
A priori, no es fácil poder modelar con certeza la forma que adopta este labio dorsal dado que su recorrido es inferior a una vuelta y, consecuentemente, las referencias sobre las que apoyarse puede que no sean suficientes. Lo que sí puede observarse es un posible crecimiento en espiral, es esperable que sea así ¿verdad?, y además que ésta sea una espiral geométrica para que el crecimiento sea gnomónico; por tanto, planteamos y establecemos esta hipótesis. Adicionalmente, por uniformidad, consideraremos que el tipo de crecimiento sea también cordobés ya que la pared dorsal en el segundo y tercer verticilo tiene esa forma (las paredes dorsales en estos verticilos eran previamente, en la vuelta anterior, sendas paredes ventrales ) y es lógico que ésta se mantenga en el primero o mejor dicho que ésa sea la tendencia original que continúa posteriormente. Es esperable y lógico que la curvatura del dorso del animal se mantenga en toda su existencia. Bajo esa consideración, lo que sí es obvio (observar la Fig. 16) que:
- La espiral que modela la pared ventral no modela al labio dorsal. Observad como la parte correspondiente a esta espiral que se ha dibujado en la imagen mediante puntos no se ajusta a dicho labio.
- El polo de la espiral del labio no puede coincidir con el polo de la espiral de la pared ventral ya que el primero ha de ubicarse en el interior del ombligo y el segundo según lo ya modelado se sitúa sobre su borde (ver Fig. 21). Estamos hablando de una distancia entre esos dos polos que estará en el entorno de un milímetro y que puede parecer una diferencia no significativa, sin embargo, como veremos, ésta es realmente la clave diferenciadora y la causa de las variaciones que se observan en el crecimiento durante el primer verticilo respecto al de los restantes. Sí, ¡así es de simple!. A nivel global, a la escala de la concha de un ejemplar maduro, no hay una diferencia visual entre ambas (ver en la esquina superior derecha en la figura 21 que la espiral ventral, en color azul, y la dorsal en rojo se "solapan"), pero a nivel local se contempla un comportamiento muy diferenciado (ver la parte central en esa figura 21), ¡esas son las maravillas de la espiral logarítmica o geométrica o "Spira mirabilis" como la bautizó Jacob Bernoulli.
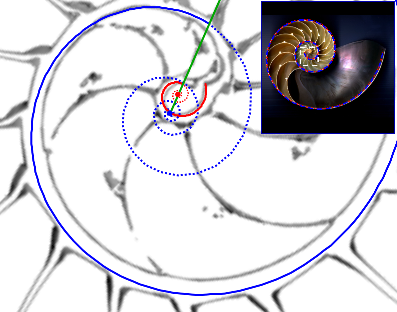
Fig. 21. Modelización de la concha en el primer verticilo. Espirales cordobesas con diferente polo. En rojo el labio dorsal y en azul la pared ventral. Diferenciado a nivel microscópico (imagen central) y "coincidente" a nivel macroscópico (esquina superior derecha).
¡Vaya! Ya lo había adelantado literariamente antes, pero ahora ¡se me hizo irresistible mostrarles gráficamente, en esta figura 21, la causa o germen que permitirá comprender mejor al Nautilus! Bien sabe quien se dedica a investigar que no puede ocultarse la satisfacción cuando en el interior de nuestra mente surge la inspiración ¡Ajá! que sublimó Martin Gardner y la que llevó a Arquimedes a correr desnudo, insconcientemente, a la par que gritaba ¡Eureka! a sus paisanos de Siracusa. Y salvando las distancias y comparaciones, estaba deseoso de que lo vieran. Pero esa inspiración inicial, que esporádicamente regalan las musas generalmente sólo a quien está trabajando, no es más que la piedra angular que asienta y modula la futura estructura global, pero habrá que seguir construyendo sobre ella en base a nuevas inspiraciones y deducciones lógicas. Así pues, hemos de asentar esa piedra base y aplicar todos los detalles detectados, analizados y descritos en la particular revisión que hemos realizado de la ontogenia biologica del Nautilus, para así llegar con paso firme a nuestro objetivo que es poder comunicarles lo que sotto voce nos cuenta el Nautilus acerca de su ser y crecer, y poder hacerle de traductor al lenguaje que muchos catalogan como el criptográfico o secreto de la Naturaleza: el matemático. Ahora sí, llegó el momento esperado, o al menos anunciado, de formular detalladamente la Ontogenia matemática del Nautilus. Gracias por ser pacientes, considero que se verán gratificados por haber llegado aquí y por continuar conmigo.
Ontogenia matemática del primer verticilo del Nautilus
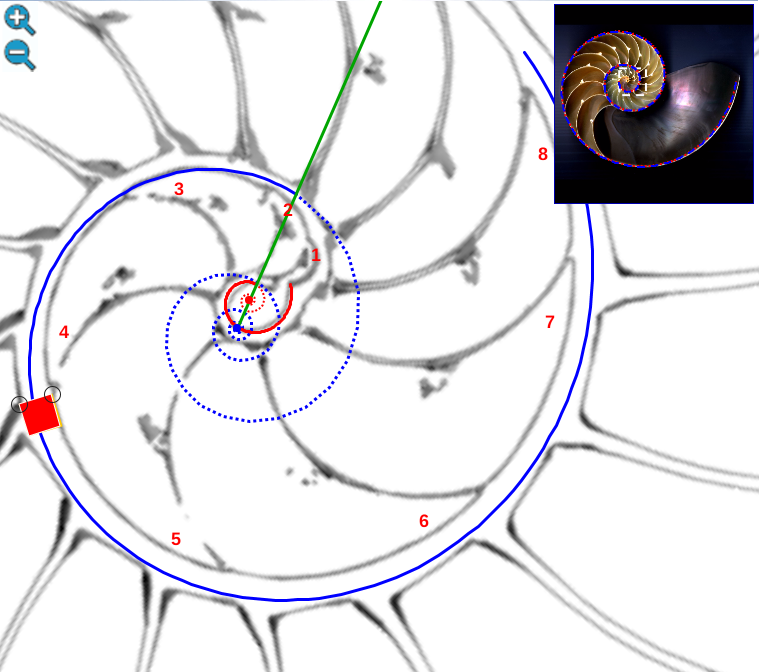
Para realizar el análisis matemático en este primer verticilo utilizaremos el siguiente recurso interactivo (escena interactiva 2):
Escena interactiva 2. Concha y sifúnculo del Nautilus en el primer verticilo.
Puede interactuar libremente con ella. Si posiciona el ratón sobre los botones tendrá una breve información o consulte las instrucciones.
Modelando el labio dorsal
Para mostrar la modelización del labio dorsal y el primer verticilo hemos elegido la concha digitalizada por el Museo de D’Arcy Thompson (Fig. 22) en la Universidad de Dundee y, en particular, los datos tridimensionales aportados por dicho museo correspondientes a esa digitalización. A partir de ellos, González-Restrepo (2019) obtuvo la sección sagital y las parasagitales (Fig. 23), así como las radiales. Realizamos esta elección porque en esta fuente gráfica el corte con los septos del primer verticilo y con los restos del canal sifuncular es más visible que los que pueden observarse en la concha que hemos utilizado, hasta ahora, como soporte de referencia para mostrar el modelo uniforme y las propiedades observadas. Aquí se puede ver que los septos del primer verticilo están incompletos y consecuentemente tampoco se visualiza el rastro del canal del sifúnculo, por tanto, no es posible modelar lo que no es visible. Esta es la razón de cambiar la referencia gráfica que utilizamos como muestra.
| Modelo tridimensional de la sección de la concha de un Nautilus pompilius. |  |
|
Fig. 22. Nautilus Shell (sectioned) by University of Dundee Museum Collections on Sketchfab |
Fig. 23. Cortes por planos paralelos al plano medio obtenidos por González-Restrepo (2019) a partir del modelo 3D del Museo Dundee. |
Partamos de que la espiral logarítmica cordobesa[1] que modela la pared ventral (representada en color azul en las imágenes y escenas) viene dada en coordenadas polares por:
![]() (1)
(1)
o en coordenadas cartesianas en función del parámetro θ :
 (2)
(2)
es decir, que ubicamos su polo en el origen de coordenadas. Adicionalmente denotaremos el ángulo inicial de la espiral como θi, y como θf el ángulo final correspondiente a la boca de la concha.
Con la notación anterior, entonces el polo de la espiral cordobesa que aproxima al labio dorsal se situaría en D(0.035, 0.080) y su ecuación paramétrica sería:
 (3)
(3)
Donde el coeficiente ![]() , verificaría que
, verificaría que ![]() .
.
De los valores posibles el que teóricamente podría ser más certero se correspondería con:
![]() (4)
(4)
pues matemáticamente representaría que el labio dorsal tiene un retraso de dos vueltas (un ángulo de retardo de 4π) respecto a la pared ventral inicial. Sería de esperar que el retraso fuera de 2π, pues ésta es la relación que acontece en los siguientes verticilos cuando la que antes era pared ventral pasa a ser la nueva pared dorsal en la siguiente vuelta (de ahí lo de 2π), pero lo esperado no es siempre lo encontrado.
Geométricamente, si el labio dorsal en el primer verticilo tuviera sólo el retardo de 2π se generaría un ombligo de mayor tamaño al que se observa en la concha del Nautilus. Esto representaría al animal un esfuerzo constructivo mayor, pues tendría que cubrir una superficie más amplia (en este caso más longitud dado que estamos trabajando con la sección) ya que este labio dorsal bordearía a la concha embrionaria. En la escena interactiva 3, espacio izquierdo, puede verse cuál sería la forma del labio dorsal, ombligo y concha en el supuesto de un retardo de 2π y en el espacio derecho el caso de retardo 4π. Entre las dos alternativas la correspondiente a 4π parece la más eficiente. Podrían plantearse retardos que no tuvieran que ser múltiplos de 2π, pero de partida ello no se observa como biológicamente adecuado, pues estaría considerándose que la parte dorsal del cuerpo del animal tendría un crecimiento descompensado, al menos en algún instante o periodo temporal, respecto a la ventral. Pudiera acontecer, pero el deslizamiento que realiza a lo largo del fragmacono al crecer, inspira más una situación más regular. Si en el periodo evolutivo acontecido en la larga historia de este animal han ocurrido retardos de otro tipo, al menos parece que la selección natural ha conducido al retardo de 4π que hemos detectado y que consideramos en nuestro modelo del primer verticilo.
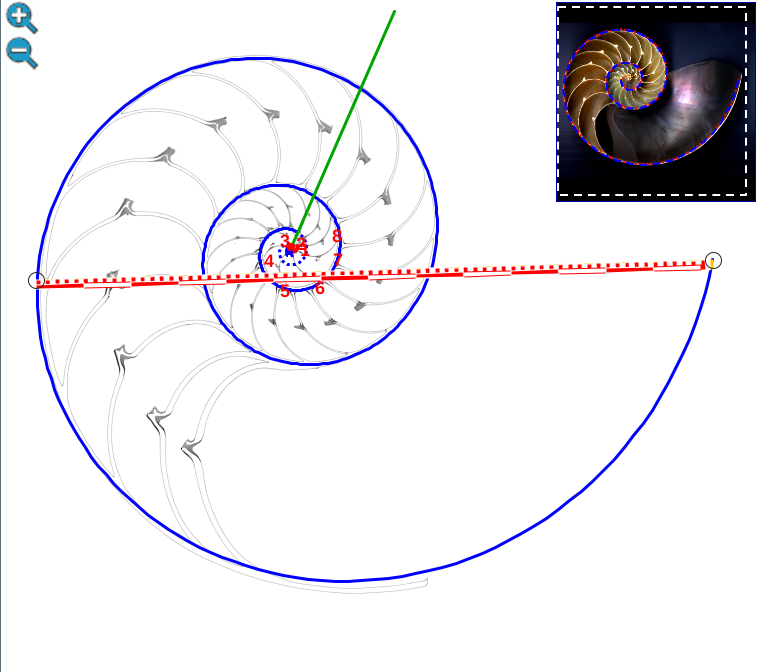
Escena interactiva 3. Modelando el labio dorsal del Nautilus en el primer verticilo.
Puede interactuar libremente con ella. Si posiciona el ratón sobre los botones tendrá una breve información o consulte las instrucciones.
Ambas espirales comparten el mismo eje polar (en color verde en la escena), es decir, el eje de cada una de ellas está incluido en la semirrecta que pasa por el polo ventral y por el polo dorsal. Adicionalmente, esta semirrecta interseca a la concha en la constricción dorsal y la ventral que son los puntos de intersección y tangencia, respectivamente, del primer septo con la pared dorsal y la ventral de dicha concha. Esto cuadra con la ontogenia biológica descrita antes y matemáticamente aporta también una pauta común de crecimiento al compartir ambas espirales el mismo parámetro angular. El ángulo inicial común para ambas espirales indicamos que lo denotaríamos como θi y entonces el recorrido total de la espiral dorsal termina en ![]() .
.
La pequeña variación existente entre los polos de ambas espirales (aproximadamente un milímetro) conduce a que el labio dorsal no finalice su recorrido siendo tangente a la concha embrionaria preseptal, sino que este labio incide e interseca a dicha concha primigenia cuando se cierra el primer verticilo y se genera el ombligo. Consecuentemente, el enlace con la parte ventral inicial no es suave, sino que se produce a través del séptimo y octavo septo. Son estos últimos septos del primer verticilo los que al apoyarse en la concha primigenia se adaptan para dar continuidad con la pared ventral inicial. El hecho de que no se produzca esa tangencialidad entre la pared ventral inicial y la finalización del labio dorsal refuerza y muestra la necesidad de considerar la existencia de dos polos diferenciados en el modelo teórico y, también, que no se pueda atribuir esta diferencia a un error en la precisión del ajuste gráfico realizado[2]. Ver detalle del modelo a nivel local en la figura 22 y a nivel global en la figura 23.
 |
 |
|
Fig. 22. Ajuste del labio dorsal (espiral en rojo) y de la pared ventral en el primer verticilo (espiral azul)). La sección gráfica de referencia se debe a González-Restrepo (2019). En rojo la regla de medida que marca 1 mm. |
Fig. 23. Ajuste global de la concha del Museo de D’Arcy Thompson, en base al corte realizado por González-Restrepo (2019). La regla de medida marca 143 mm. |
El canal sifuncular en el primer verticilo
En nuestro primigenio modelo uniforme detectamos que el canal sifuncular actuaba como el eje del fragmacono, es decir, que es equidistante de la pared ventral y dorsal. En este supuesto uniforme ambas paredes describen sendas espirales cordobesas con igual polo, de manera que si la ecuación en coordenadas polares de la primera es ![]() , la de la segunda sería
, la de la segunda sería ![]() (realmente es la misma espiral, pero la segunda con un ángulo de retardo de 2π o coeficiente 0,3431...) y, entonces, el sifúnculo tiene de ecuación
(realmente es la misma espiral, pero la segunda con un ángulo de retardo de 2π o coeficiente 0,3431...) y, entonces, el sifúnculo tiene de ecuación
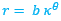 (5)
(5)
donde 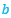 toma el valor medio de los respectivos coeficientes 1 y
toma el valor medio de los respectivos coeficientes 1 y 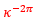 de las espirales anteriores, es decir,
de las espirales anteriores, es decir,
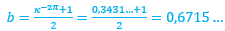 (6)
(6)
lo que se corresponde con una espiral cordobesa con un ángulo de retardo de ![]() . Pero, ya comprobamos que el ajuste anterior es adecuado en el segundo y tercer verticilo, pero no así para el primero. No obstante, ahora sí, estamos en condiciones de determinar cómo queda modelado el sifúnculo en esta primera vuelta.
. Pero, ya comprobamos que el ajuste anterior es adecuado en el segundo y tercer verticilo, pero no así para el primero. No obstante, ahora sí, estamos en condiciones de determinar cómo queda modelado el sifúnculo en esta primera vuelta.
El fragmacono en el primer verticilo está delimitado por dos espirales cordobesas cuyas ecuaciones son (2) y (3), ambas con un ángulo polar común, pero con distinto polo. Si consideramos que el sifúnculo en este verticilo tiene un comportamiento similar a los dos posteriores, entonces éste sería el eje de ese fragmacono y, para determinarlo, procedamos a determinar el lugar geométrico de los puntos del plano que equidistan de ambas y obtendremos que viene determinado por la ecuación:
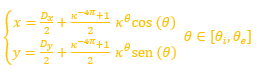 (7)
(7)
que se corresponde con una espiral cordobesa, cuyo polo![]() es el punto medio de segmento determinado por el polo de la pared dorsal
es el punto medio de segmento determinado por el polo de la pared dorsal ![]() y el de la ventral
y el de la ventral ![]() , y su coeficiente es la media aritmética de los coeficientes respectivos[3]
, y su coeficiente es la media aritmética de los coeficientes respectivos[3]
![]() (8)
(8)
Al representar gráficamente esta espiral (ver en la imagen derecha de la fig. 24 la línea discontinua en color naranja o interactuar en la escena interactiva 3) observamos que realmente no modela adecuadamente a dicho sifúnculo en ese verticilo. Sin embargo, si observamos en esa figura 24 la espiral trazada en una línea continua, también de color naranja, ésta sí se ajusta al los restos del canal sifuncular.
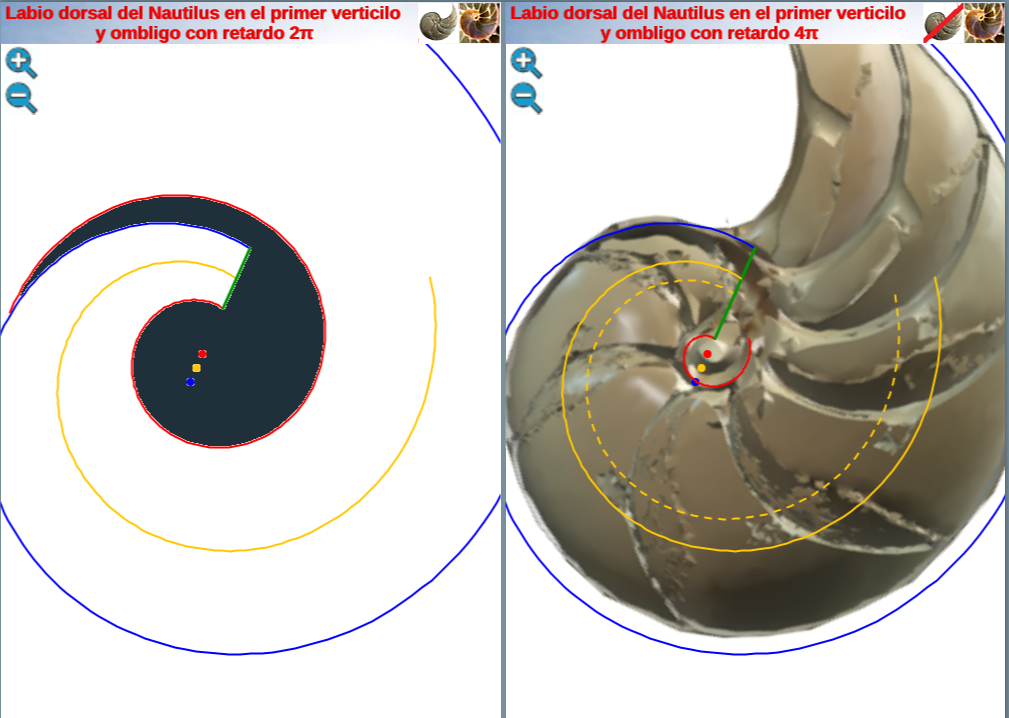
Fig. 24. En la imagen de la izquierda y de la derecha eje del fragmacono en el primer verticilo cuando el labio dorsal tiene un retardo de 2π (espiral continua de color naranja). A la derecha en línea discontinua eje del fragmacono en el primer verticilo cuando el labio dorsal tiene un retardo de 4π. La primera se ajusta al canal sifuncular.
La ecuación de esa espiral y sifúnculo es la siguiente:
 (9)
(9)
donde
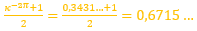 (10)
(10)
y se correspondería con el eje del fragmacono, pero en el caso de que el labio dorsal tuviera sólo un retardo de 2π (ver imagen izquierda en la figura 24 o interactuar en la escena 3). Ese coeficiente o retardo en el sifúnculo es el mismo que acontece en el segundo y tercer verticilo, pero aquí el polo es diferente. En la figura 25 podemos ver una comparativa de la espiral sifuncular aquí determinada (en color naranja) y la del sifúnculo en el modelo uniforme (en turquesa).
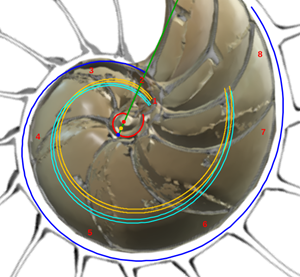
Fig. 25. Comparativa de la espiral sifuncular del primer verticilo (naranja) y la del sifúnculo en el modelo uniforme (turquesa).
Así pues, la causa de que el sifúnculo tenga un comportamiento diferenciado en el primer verticilo se centra en el hecho de que el labio dorsal y la pared ventral tienen diferente polo. La explicación de cuál es la causa de su desviación respecto al eje del fragmacono queda abierta...
Pienso que hemos dado un gran paso en la comprensión y explicación de la ontogenia matemática del Nautilus. En ella, sigue imperando su carácter netamente cordobés, introduciendo bonitos matices matematicos que explican los matices biológicos observados. Pero D. Antonio Machado nos indicó que se hace camino al andar y para ello, después del primer paso dado, han de llegar otros y en nuestro estudio, ahora, tendremos que centrarnos especialmente con las paredes septales y, ¿por qué no?, quizás tratar de localizar cuál es el germen de la esencia cordobesa de este entrañable, al menos para mí, animalito. Así pues, tendré que preparar un nuevo artículo. ¿Será el último de esta serie o no será? El tiempo y la inspiración lo dirá... Les estaré esperando por aquí... ¡pronto!
[1] Para saber por qué esa espiral es una espiral cordobesa, por qué la razón de que la base de esa exponencial sea κ = 1,18558… y su significado, puede consúltarse: Galo et al (2016).
[2] De nuevo, hemos de recordar que el paso de la yocto-yotta realidad al modelo teórico requiere de la consideración de cierta tolerancia, pero que hay que saber distinguirla de la existencia de una diferencia que provoque cambios en el modelo. Si la toma en consideración de esa tolerancia no cambia el modelo podremos aceptarla y admitirla, pero habrá que evitarla cuando se provoque un cambio en él. Así pues, en este caso es obligado considerar que los polos son diferentes. Por el contrario, por ejemplo, en las espirales que hemos considerado para ajustar el perfil del labio dorsal y de la pared ventral, en especial ésta, podría aducirse que necesitarían una mejora en su ajuste a la imagen de la concha en ese primer verticilo, pero realmente no es necesario ya que tal y como se ha considerado es suficientemente adecuado para determinar el modelo global que caracteriza al genus Nautilus. Esto ya lo mostramos en la fig. 21, y lo detallamos ahora, pues al observar el ajuste de la pared ventral (espiral azul) en el primer verticilo y su ajuste global en la toda la concha, se marca de manera evidente que la tolerancia considerada es admisible para determinar el modelo a nivel tanto local como global (el grosor de la pared es inferior a un mm, lo puede medir en la escena y el ejemplar considerado tiene unos 143 mm en su etapa adulta, figuras 22 y 23) . Sin embargo, si modificásemos la posición del polo de la pared dorsal, es decir, ignorásemos la pequeña distancia que existe entre ambos polos obtendríamos variaciones no admisibles a nivel local, es decir, cambiaría el modelo y no se ajustaría a la ontogenia del Nautilus.
[3] En general puede comprobarse que dadas dos espirales logarítmicas con igual base, con el mismo ángulo polar y con diferentes polos; si se halla el lugar geométrico de los puntos de plano obtenidos mediante una combinación lineal de las mismas se obtiene que es una espiral logarítmica del mismo tipo, es decir, de igual base y ángulo polar, y con polo la combinación lineal de los polos y coeficiente la combinación de los coeficientes.
Bibliografía
Galo J.R., Cabezudo A. y Fernández I.(2016 a) : Sobre la forma y crecimiento cordobés del Nautilus Pompilius. Epsilon, 2016, Vol. 33 (3), nº 94.
González-Restrepo, F. (2019): Cortes del Nautilus a partir de la digitalización 3D del museo Dundee. Red Descartes Colombia.
Landman, N. H., Arnold, J. M. and Mutvei, H. 1989: Description of the embryonic shell of Nautilus belauensis. American Museum Novitates, no. 2960, p. 1–16.
Mutvei, H. and Doguzhaeva, L. 1997: Shell ultrastructure and ontogenetic growth in Nautilus pompilius L. (Mollusca: Cephalopoda). Palaeontographica Abteilung A Palaeozoologie–Stratigraphie, vol. 246, p. 33–52.

Este obra está bajo una licencia de Creative Commons Reconocimiento-NoComercial-SinObraDerivada 4.0 Internacional

 CONTACTO
CONTACTO


