OBJETIVOS
- Determinar un tipo que refleje la forma de la sección frontal del Nautilus necesaria para modelar la concha tridimensional del Nautilus.
- Mostrar las dificultades encontradas en la determinación de ese tipo.
INTRODUCCIÓN
En esta miscelánea se aborda la comparación de diferentes secciones frontales del Nautilus obtenidas en varios artículos de Tanabe et al. de 1982 a 1995 en los que desglosan su investigación sobre la morfología y biometría de este cefalópodo, pero hay más artículos donde estos autores han divulgado su investigación en estos asuntos y en los que he accedido a sus planteamientos científicos. También se incluirán las secciones de la concha digitalizada del Museo de D'Arcy Thompsosn realizadas por González-Restrepo en 2019 y también recursos propios (Galo 2023).
Los artículos son:
- Galo J.R., Cabezudo A. y Fernández I.(2016) : Sobre la forma y crecimiento cordobés del Nautilus Pompilius. Epsilon, 2016, Vol. 33 (3), nº 94.
- Galo J.R., (2023) : Modelo matemático ontogénico del Nautilus. Red Educativa Digital Descartes.
- González-Restrepo, F. (2019): Cortes del Nautilus a partir de la digitalización 3D del museo Dundee. Red Descartes Colombia.
- Hayasaka, Shozo, Tanabe, Kazushige et al. (1982). Field study on the habitat of Nautilus in the environs of Cebu and Negros Islands, the Philippines. Mem. Kagoshima Univ. Res. Center S. Pac., Vol. 3, No. 1. 1982 p. 67-137.
- Tanabe, Kazushige(1985). Record of Trapping Experiment. In: Hayasaka, S. (ed.) Marine ecological studies on the habitat of Nautilus pompilius in the environs of Viti Levu, Fiji. Kagoshima University, Research Center for the South Pacific, Occasional Papers, 4, 10-17..
- Tanabe, Kazushige(1988). Record of Trapping Experiment. Kagoshima University, Research Center for the South Pacific, Occasional Papers, No. 15, p.5-15.
- Tanabe, Kazushige & Hayasaka, Shozo & Tsukahara, Junzo. (1985). Morphological Analysis of Nautilus pompilius. Kagoshima Univ. Res. Center S. Pac..Occasional Papers, No. 4. p. 38-49.
- Tanabe, Kazushige & Tsukahara, Junzo. (1987). Biometric Analysis of Nautilus pompilius from the Philippines and the Fiji Islands. In book: Nautilus: The Biology and Paleobiology of a Living Fossil. Chapter: 7. Publisher: Plenum Publishing Corporation. Editors: W. Bruce Saunders and Neil H. Landman.
- Tanabe, Kazushige & Tsukahara, Jyunzo & Hayasaka, Shozo. (1990). Comparative morphology of living Nautilus (Cephalopoda) from the Philippines, Fiji and Palau. Malacologia 31(2):297-312.
- Tanabe, Kazushige & Tsukahara, Jyunzo & Shinomiya, Akihiko & Oki, Kimihiko. (1991). Notes on Nautilus Pompilius captured from Port Moresby area, Papua New Guinea. Kagoshima Univ. Res. Center S. Pac., Occasional Papers, No. 21. p. 33-38.
- Tanabe, Kazushige & Tsukahara, Junzo. (1995). Morphological Analysis of Living Nautilus from Palau. Kagoshima Univ. Res. Center S. Pac., Occasional Papers, No. 27. p. 41-55.
El conocimiento de cómo es la sección fromtal del Nautilus es algo básico e imprescindible para poder abordar la modelación tridimensional del Nautilus y de ahí nuestro foco de interés. Según Raup, D. M. (1966) en el artículo "Analysis of Shell Coiling: General Problems." toda concha puede modelarse en base a cuatro parámetros esenciales: curva generatriz o sección transversal, la distancia o posición relativa de la curva generatriz respecto al eje de giro, factor de crecimiento y la traslación en la dirección del eje de giro (sólo para conchas turbinadas, lo cual no acontece en el Nautilus ya que es discoidal).
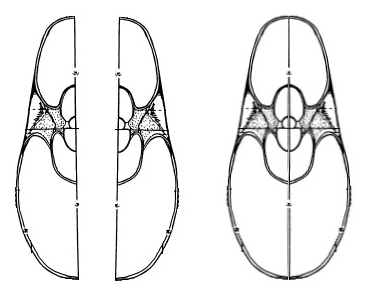
La sección del Nautilus con la que comencé a trabajar es la reflejada enTanabe, Hayasaka & Tsukahara (1985) y que se muestra en la parte derecha de la imagen siguiente:
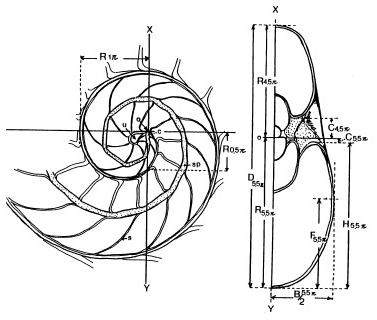
Figura 1. Sección sagital de la concha nepiónica y sección frontal (Tanabe et al., 1985).
y a partir de ella editando la imagen se puede conformar la sección completa:
Figura 2. Sección frontal (Tanabe et al., 1985).
Pero desafortunadamente, para mí, la imagen de la sección frontal reflejada en Tanabe et al. (1985) no puede ser correcta. En mi opinión, cuando editaron ese artículo, esta imagen debió de sufrir un cambio no proporcional quedando deformada, pues después de tomarla como tipo para la modelación y trabajar con ella detecté, como detallaré posteriormente, que no seguía la proporcionalidad cordobesa que guía y es intrínseca al Nautilus (Galo et al. 2016 y Galo 2023).
Análisis de los datos y de la sección frontal aportados por Tanabe et al. (1985)
Era necesario abordar un contraste entre esa imagen (figura 1) y los datos aportados por Tanabe et al. en el mismo artículo de 1985 y en otros artículos que indicamos después, y así poder verificar la certeza de la incongruencia detectada y abordar la elección de un neotipo para la sección frontal del Nautilus. Por ello destaquemos:
- La imagen citada (tanto de la sección sagital de la concha nepiónica, como de la sección frontal) aparece en los artículos Tanabe et al. (1985) y en Tanabe & Tsukahara (1987) y los datos numéricos relativos a los radios vectores de la concha de 28 ejemplares tomados cada π/4 radianes se reflejan también en dicho artículo de 1985.
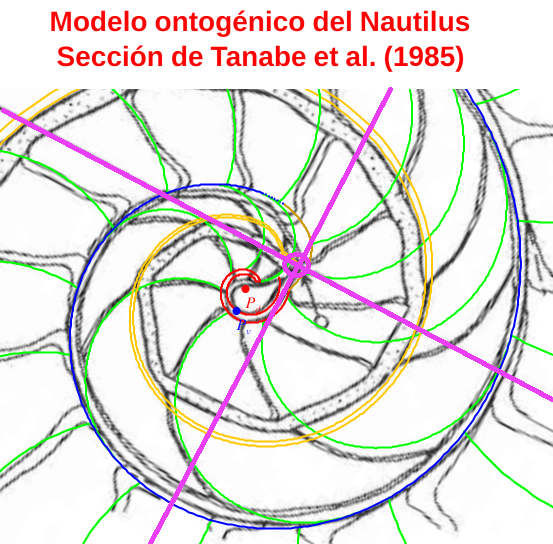
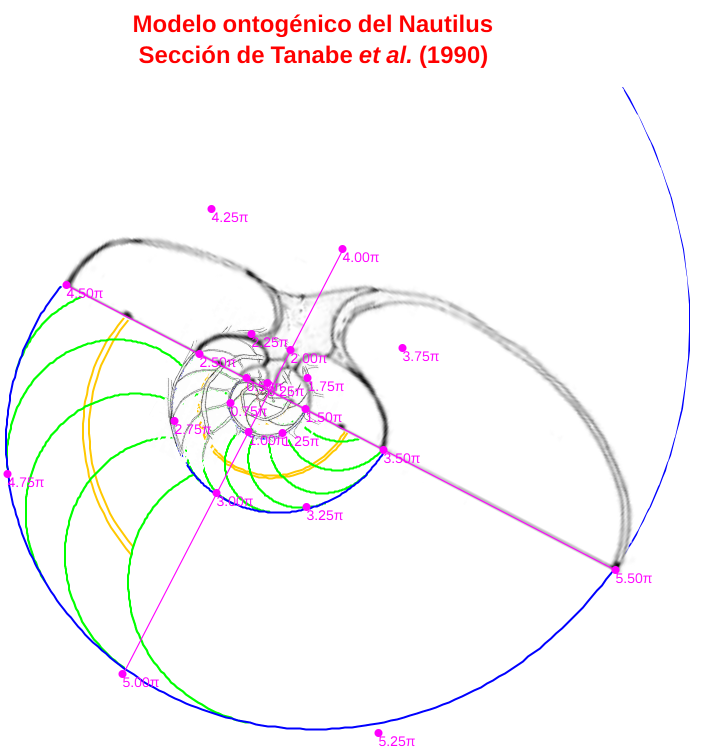
- Para la medida de los radios vectores estos autores consideran que el polo de la espiral que determina la sección sagital de la concha del Nautilus ha de ubicarse en el inicio del sifúnculo (p. 39, p. 42 y p. 43 en Tanabe et al., 1985). Adoptan este criterio e indican en la p. 42 de ese artículo que no coincide con lo realizado por otros autores que sitúan el polo en el centro del ombligo. Tampoco, indico ahora yo, coincide con lo realizado en el modelo cordobés uniforme (Galo et al., 2016) ni en el ontogénico (Galo, 2023) donde el polo de la pared ventral es exterior a la concha embrionaria inicial que con el primer septo conforma la primera cámara, y no coincide tampoco con el centro del ombligo. Puede verse la diferencia entre estos puntos en la siguiente imagen (figura 3), donde se refleja en color azul el polo ventral (Pv), en rojo el polo de la espiral dorsal (Pd) que aquí no interviene, el polo que denominaremos como polo de Tanabe resaltado con una circunferencia en color magenta, señalado también por una flecha en la imagen original, y en el mismo color los ejes de cordenadas que consideraron estos autores, donde en esta imagen el eje polar r = 0 es la semirrecta con pendiente positiva en sentido creciente y la medición de ángulos es en sentido positivo, es decir, con crecimiento levógiro (la imagen de Tanabe, ver figura 1, está en sentido dextrógiro y el eje polar es el horizontal en sentido hacia la derecha, aquí la hemos transformado isométricamente para superponerla adecuadamente al modelo ontogénico).
Figura 3. Ubicación de los polos y ejes en la Sección sagital de Tanabe et al., 1985.
- La sección frontal —p. 107 en Tanabe et al., 1987— indican que la realizan tomando la media concha, a partir de la que se ha obtenido la sección sagital, y volviéndola a cortar ahora perpendicularmente al plano sagital y siguiendo el eje longitudinal a lo largo del sifúnculo (marcado por el segmento XY indicado en ambas imágenes, izquierda y derecha, reflejadas en la figura 1). Por ello, en dicha sección frontal se observa una mitad de los cortes frontales de la concha que en la imagen se corresponden según los autores con los ángulos 0,5π, 1,5π, 2,5π, 3,5π, 4,5π y 5,5π (por proporcionalidad y en un supuesto crecimiento gnomónico uniforme podría asociarse con cualquier serie de valores en progresión aritmética de diferencia π).
- En la hoja de cálculo datos del Nautilus según Tanabe et al. (1985), machos y hembras, puede verse los datos aportados por estos autores donde he incluido también el análisis del factor crecimiento obtenido a partir de ellos. También se tienen los datos diferenciando por sexo: "Machos" y "Hembras", si bien el número de hembras en mucho menor que el de machos. El error relativo en el factor de crecimiento puede consultarse en la hoja "Error relativo global, machos y hembras". Todos estos datos son relativos al polo y ejes considerados por estos autores (figura 3).
Si consideramos el polo ventral indicado por Galo (2023) y, a partir de los datos anteriores, calculamos el radio vector que corresponde en este caso a todos esos puntos obtenemos la tabla "Datos del Nautilus desde el polo ventral (Galo, 2023)" donde también incluimos el análisis del factor de crecimiento, teniendo también "Error relativo global desde el polo ventral, machos y hembras".
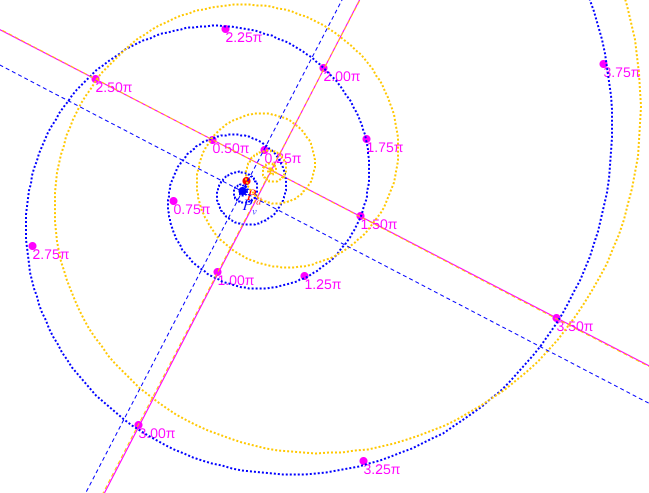
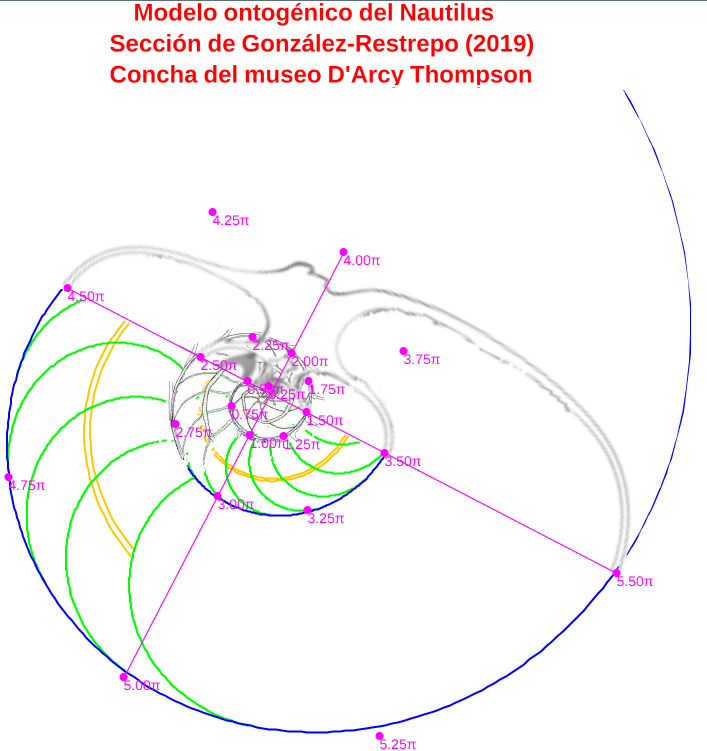
En un análisis de los datos anteriores se concluye que el modelo de crecimiento según una espiral logarítmica, que es lo esperado, y en particular el que ésta sea una espiral cordobesa, se ajusta mejor en el caso de considerar el polo ventral indicado por Galo (2023) frente al polo considerado por Tanabe et al. (1985). En la imagen de la figura 4 podemos observar los datos de Tanabe et al. (1985) dibujados (puntos magenta) de manera superpuesta sobre el modelo ontogénico (Galo, 2023) y cómo estos se ajustan globalmente de forma adecuada a este modelo teórico. Puede también interactuar con la escena " Secciones frontales y modelo ontogénico del Nautilus" de donde hemos obtenido esta imagen y las siguientes.
Figura 4. Datos de Tanabe et al. (1985) dibujados sobre el modelo ontogénico (Galo, 2023).
Y en la imagen de la figura 5 comprobamos la diferencia entre la espiral cordobesa de polo el elegido por Tanabe et al. (1985) y que pasa por el punto que define el radio vector 0,5π , dibujada en color naranja, y la espiral cordobesa de polo el indicado por Galo (2023) y que pasa por ese mismo punto, dibujada en color azul.
Figura 5. Sistema de referencia y datos de Tanabe et al. (1985) —en color naranja— y sistema de referencia de Galo (2023) —en color azul—.
Adicionalmente, si ajustamos la imagen de la sección frontal de Tanabe et al. (1985), reflejada en la figura 1 y 2, podemos observar cómo ésta no se corresponde con la datos de estos autores, ni con el modelo de crecimiento cordobés esperado (ver figura 6).
Figura 6. Sección frontal de Tanabe et al. (1985) sobre los datos de estos autores y sobre el modelo ontogénico de Galo (2023).
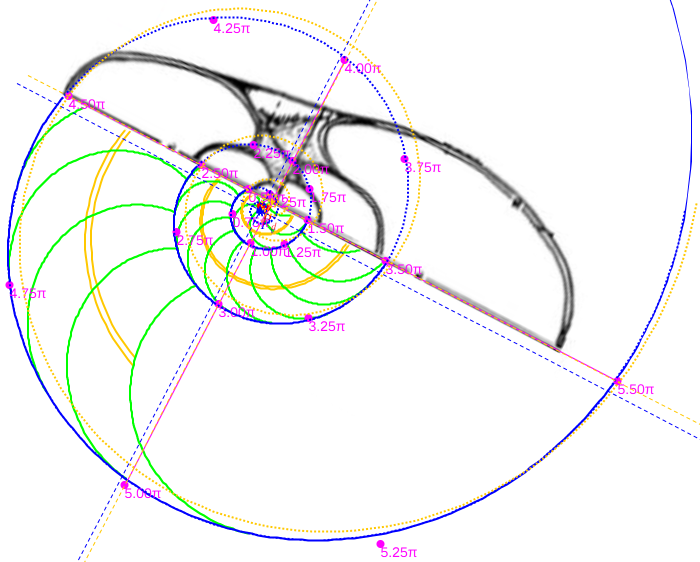
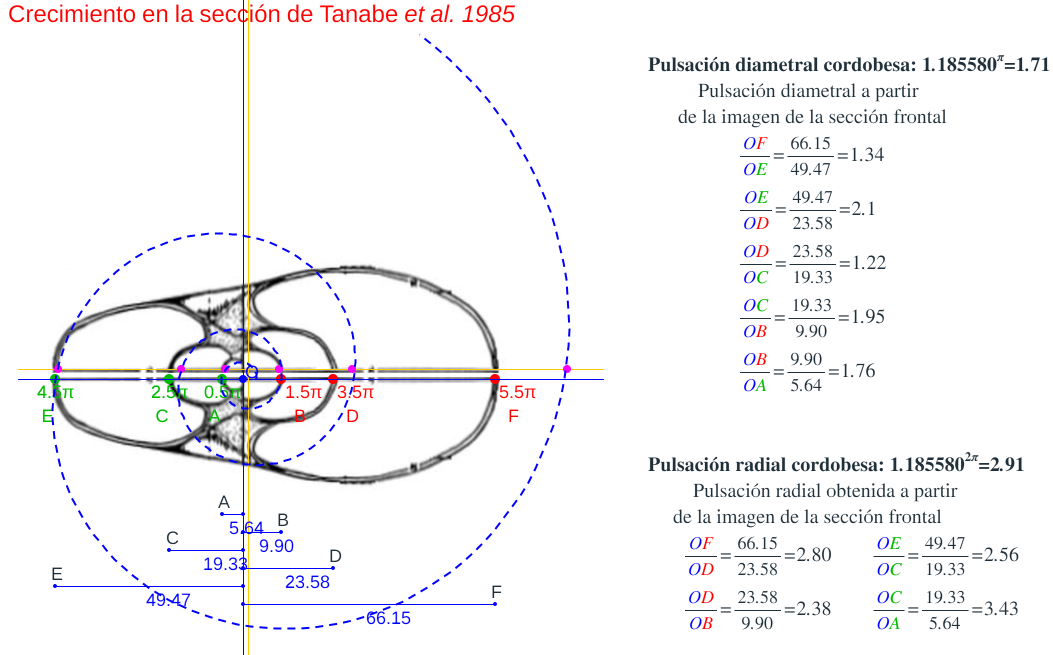
Es más, en la siguiente imagen (figura 7), en color azul, se refleja la espiral cordobesa que pasa por los puntos dados por Tanabe et al. (1985) para los ángulos 0,5π, 1,5π, 2,5π, 3,5π, 4,5π y 5,5π, dibujados de nuevo en color magenta, y que deberían de coincidir con las pulsaciones radiales de la sección frontal, pero claramente no coinciden. Si obtenemos los puntos análogos a los anteriores pero a partir de la imagen de la seción frontal, etiquetados de A a E en esa imagen, vemos que esa espiral pasa por A, B y E, pero no pasa por los puntos C, D y F, es decir, hay una discrepancia entre los datos numéricos y la imagen de la sección frontal aportados por estos autores. Así pues, si esa imagen se correspondiera con la sección correcta obtendríamos que el factor de crecimiento diametral y radial no diferiría de sendas razones correspondientes al crecimiento cordobés, pero según se refleja en las proporciones calculadas a la derecha de esa figura la diferencia es evidente. En la escena "Crecimiento en algunas secciones frontales del Nautilus" pueden verse estos detalles.
Figura 7. Crecimiento la sección frontal del Nautilus aportada por Tanabe et al. (1985).
Así pues, como había indicado con antelación, podemos concluir que la sección frontal reflejada en la figura 2 no puede corresponderse con la sección del Nautilus. Se confirma que el problema se centra en la imagen deformada y que los datos numéricos se ajustan globalmente a la proporción cordobesa esperada sin más que considerar el polo ventral indicado por Galo (2016 y 2023). Es necesario la búsqueda de otra sección frontal que nos permita abordar nuestro diseño tridimensional.
Pero previamente, veamos que las dos secciones con las que trabajaremos posteriormente, sí se ajustan a los datos de Tanabe et al. (1985) y al modelo ontogénico de Galo (2023).
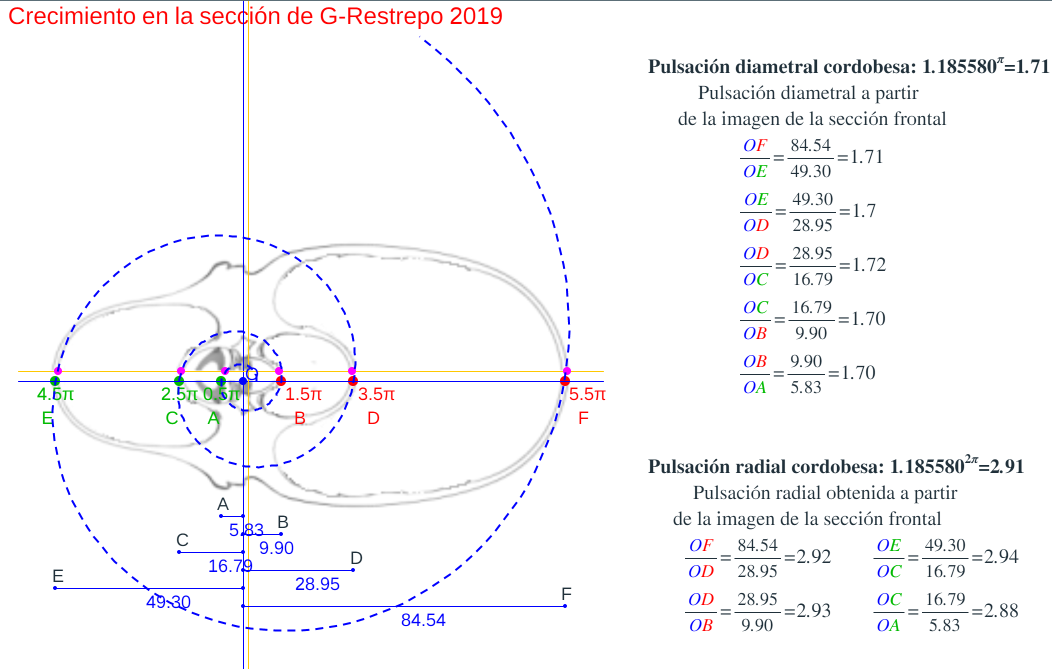
- Consideremos el corte o sección frontal que Gonzalez-Restrepo (2019) hizo a la digitalización de la concha del Nautilus del museo D'Arcy Thompson de la Universidad de Dundee y en ella podemos observar que sí se verifica la proporcionalidad y crecimiento cordobés, según se refleja en las figuras 8 y 9 y también accediendo a las escenas antes enlazadas: "Secciones frontales y modelo ontogénico" y "Crecimiento en algunas secciones frontales del Nautilus", y eligiendo la sección G_Restrepo (2019).
Figura 8. Crecimiento la sección frontal del Nautilus obtenida por G_Restrepo (2019).
Figura 9. Sección frontal de G_Restrepo (2019) sobre los datos de Tanabe et al. (1985) y sobre el modelo ontogénico de Galo (2023). .
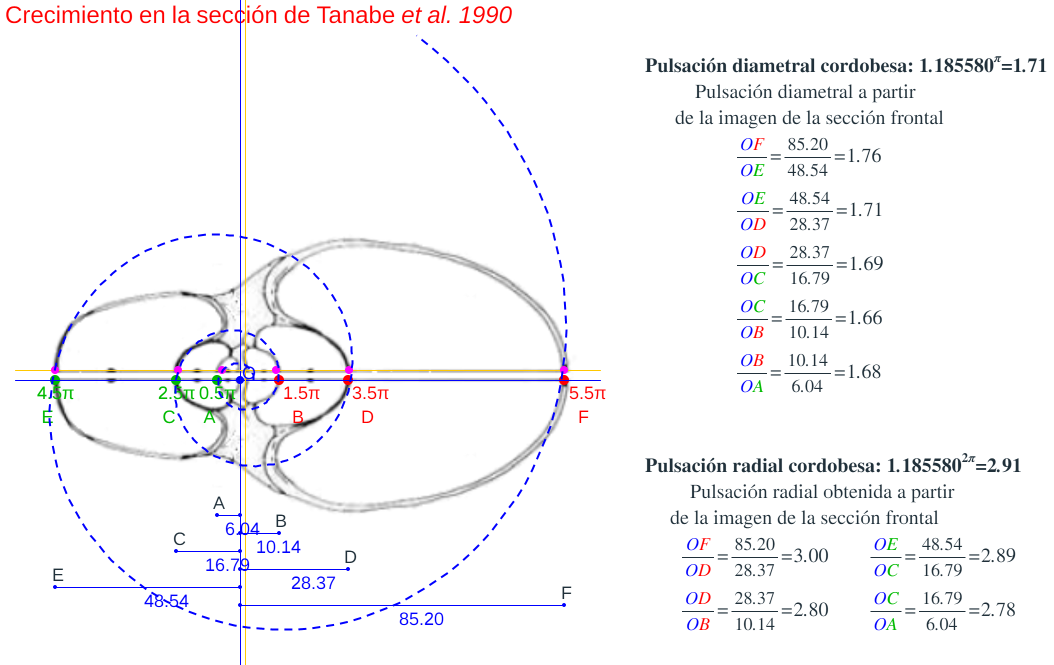
- Y lo mismo acontececon la sección de Tanabe et al. (1990) que consideraremos después y que reflejamos en las figuras 10 y 11 y en sendas escenas "Secciones frontales y modelo ontogénico" y "Crecimiento en algunas secciones frontales del Nautilus", eligiendo la sección "Tanabe 1990".
Figura 10. Crecimiento la sección frontal del Nautilus de Tanabe et al. (1990).
Figura 11. Sección frontal de Tanabe et al. (1990) sobre los datos de Tanabe et al. (1985) y sobre el modelo ontogénico de Galo (2023). .
Búsqueda de un tipo (holotipo) para la sección frontal del Nautilus
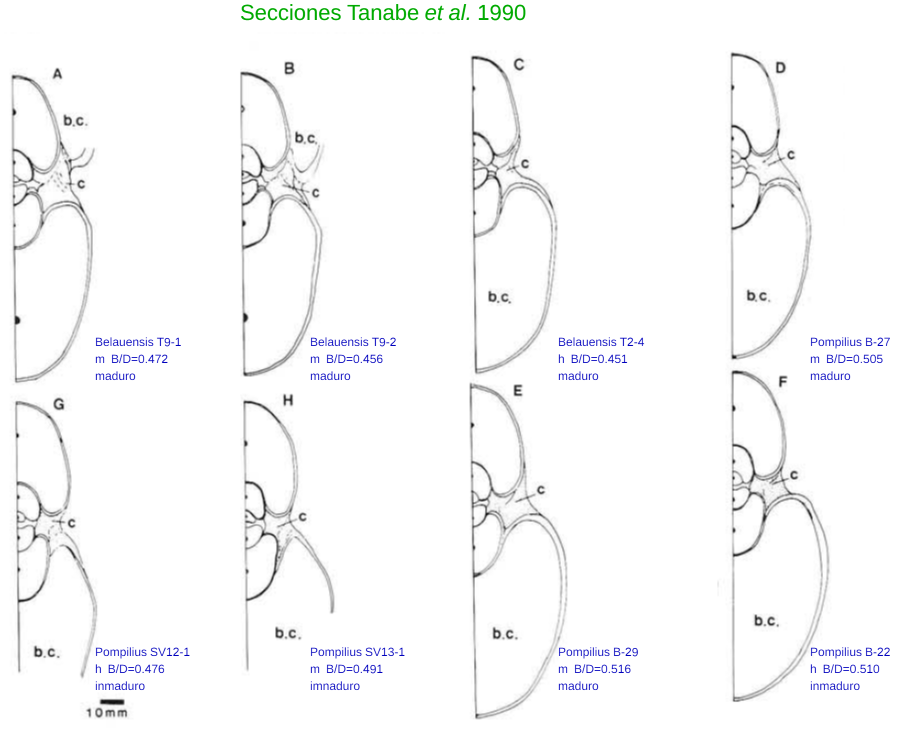
Consecuentemente se detecta la necesidad de encontrar un tipo para la sección frontal del Nautilus y si bien el corte de la concha digitalizada del Museo de D'Arcy Thompson pudiera ser considerada como tal, preferí localizar otros prototipos externos o no ligados directamente con mi investigación (Gónzález Restrepo hizo esos cortes a petición mía) y los encontré en los mismos autores (Tanabe, Tsukahara y Hayasaka) en la página 302 de su artículo de 1990 " Comparative Morphology of living Nautilus (Cephalopoda) from the Philipines, Fiji and Palau" (Tanabe et al.,1990).
En la imagen se reflejan los ejemplares incluidos en dicho artículo indicando los códigos identificadores aportados por los autores, el sexo (m-> macho, h-> hembra) y la proporción entre el ancho del verticilo de la concha (B: whorl breadth) y el diametro (D: Shell diameter) de ella (B/D). Todos los datos asociados a estos ejemplares y otros adicionales más pueden consultarse:
- Para los ejemplares A, B y C en Tanabe y Tsukahara (1995), ejemplares de Nautilus Belauensis (muy parecido al pompilius, pero de mayor tamaño, y con el que comparte el tener el ombligo cerrado y cubierto con una callosidad). Los datos usados los he reflejado también en esta página.
- Para los ejemplares D, E y F en Hayasaka et al. (1982), ejemplares de Nautilus Pompilius. Los datos también pueden consultarse en esta otra página.
- Para los ejemplares de Nautilus pompilius G y H en Tanabe (1985), que pueden completarse con más ejemplares de pompilius en Tanabe (1988).
Para la elección del tipo tendremos en consideración que:
En base a ello consideraré como tipo de la sección del Nautilus a la etiquetada como D, que a partir de ahora identifaré como tipo Tanabe (1990):
- Las secciones G y H están incompletas en la imagen, luego no podemos considerarlas para este fin, hay que excluirlas.
- En Hayasaka et al. (1982, p101) se indica que la proporción B/D no tiene variación significativa dependiente del sexo y también indica que la media aritmética de la proporción B/D en los 52 ejemplares analizados en ese estudio es de 0,4964 para los machos, 0,4926 para las hembras y 0,4938 para todos. Ver la tabla 11 de ese artículo que reflejamos a continuación:
Por tanto, de los ejemplares D, E y F el candidato a elegir como tipo para el Nautilus Pompilius sería el D por estar más cercano a la media indicada respecto a su sexo. La diferencia respecto a la media de machos es 0,505-0,4964 = 0,0086, y con la media global 0,505-0,4938 = 0,0112
- Para los ejemplares de Belauensis la media global de la proporción B/D es 0,4771, para los machos 0,4836 y para las hembras 0,4655 (consultad los cálculos obtenidos a final de la hoja de datos) y, por tanto, el candidato a elegir como tipo para el Nauitlus Belauensis sería el A. En este caso la diferencia respecto a la media de machos 0,472-0,4836 = - 0,0116 y respecto a la media global 0,472-0,4771 = -0,0051.
Previamente en esta discusión hemos comprobado (figuras 10 y 11) que esa sección Tanabe (1990) se ajusta adecuadamente al modelo ontogénico y a los datos numéricos globales de Tanabe et al. (1985).
INSTRUCCIONES
En la escena podemos distinguir dos zonas que hemos separado por una raya vertical.En la parte inferior se dispone de dos controles tipo pulsador:
- A la izquierda se han fijado las ocho secciones del Nautilus (tres del Nautilus Belauensis y cinco del Nautilus Pompilius) procedentes del artículo de Tanabe et al. de 1990.
- A la derecha está fijada también la sección del Nautilus reflejada en el artículo de Tanabe et al. de 1985 y el corte de G-Restrepo (2019) de la concha digitalizada del museo D'Arcy Thompson. Y, adicionalmente, se incluye la sección de Tanabe et al. (1990) etiquetada como D (nuestro tipo elegido) y que puede desplazarse con el control gráfico (punto de color amarillo).
En esas secciones y mediante esas comparaciones puede intuirse el caracter isomorfo de la especie Nautilus, con el margen obvio de las particularidades de cada individuo.
- "escala la sección" que permite modificar la escala del tipo seleccionado y compararlo con el resto de secciones ahí reflejadas.
- "sección simétrica" que dibuja la sección simétrica a la considerada inicialmente y así efectuar la comparación mediante superposición con las otras
Realizado en el año del vigésimo quinto aniversario del Proyecto Descartes